met genexpressie bedoelen we de transcriptie van een gen in mRNA en de daaropvolgende vertaling in eiwit. De uitdrukking van het gen wordt hoofdzakelijk gecontroleerd op het niveau van transcriptie, grotendeels als resultaat van band van proteã nen aan specifieke plaatsen op DNA. In 1965 deelden Francois Jacob, Jacques Monod en Andre Lwoff de Nobelprijs voor de geneeskunde voor hun werk dat het idee ondersteunt dat de controle van enzymniveaus in cellen wordt gereguleerd door transcriptie van DNA., komt door Verordening van transcriptie voor, die of kan worden veroorzaakt of worden onderdrukt. Deze onderzoekers stelden voor dat de productie van het enzym wordt gecontroleerd door een “operon”, dat bestaat uit een reeks verwante genen op het chromosoom bestaande uit een operator, een promotor, een regulatorgen en structurele genen.
- De structuurgenen bevatten de code voor de eiwitproducten die moeten worden geproduceerd. De regelgeving van eiwitproductie wordt grotendeels bereikt door toegang van de polymerase van RNA tot het structurele gen te moduleren dat wordt getranscribeerd.,
- het promotergen codeert niets; het is gewoon een DNA-sequentie die de initiële bindingsplaats is voor RNA-polymerase.
- het operatorgen codeert ook niet; het is slechts een DNA-sequentie die de bindingsplaats is voor de repressor.
- de regulatorgencodes voor de synthese van een onderdrukkingsmolecuul dat bindt aan de operator en de transcriptie van de structurele genen van RNA-polymerase blokkeert.
het operatorgen is de sequentie van niet-transcribeerbaar DNA dat de bindingsplaats van de onderdrukker is., Er is ook een reguleringsgen, dat codeert voor de synthese van een onderdrukkingsmolecuul hat bindt aan de operator
- voorbeeld van induceerbare transcriptie: de bacterie E. coli heeft drie genen die coderen voor enzymen die het mogelijk maken lactose (een suiker in melk) te splitsen en te metaboliseren. De promotor is de plaats op DNA waar de polymerase van RNA bindt om transcriptie in werking te stellen. Echter, de enzymen zijn meestal aanwezig in zeer lage concentraties, omdat hun transcriptie wordt geremd door een onderdrukt eiwit geproduceerd door een regulator gen (zie het bovenste gedeelte van de figuur hieronder)., Het repressoreiwit bindt aan de plaats van de operator en remt de transcriptie. Nochtans, als lactose in het milieu aanwezig is, kan het aan de repressorproteã ne binden en het inactiveren, effectief het verwijderen van de blokkade en het toelaten transcriptie van boodschappersRNA nodig voor synthese van deze genen (lager gedeelte van het Cijfer hieronder).

- voorbeeld van onderdrukbare transcriptie: E. coli heeft het aminozuur tryptofaan nodig en het DNA in E. coli heeft ook genen voor de synthese ervan., Deze genen transcriberen over het algemeen continu omdat de bacterie tryptofaan nodig heeft. Als tryptofaanconcentraties echter hoog zijn, wordt de transcriptie onderdrukt (uitgeschakeld) door binding aan een repressoreiwit en activatie zoals hieronder weergegeven.
![]()
controle van genexpressie in eukaryoten
eukaryotische cellen hebben vergelijkbare mechanismen voor controle van genexpressie, maar ze zijn complexer., Overweeg, bijvoorbeeld, dat prokaryotic cellen van een bepaalde species allemaal hetzelfde zijn, maar de meeste eukaryotes zijn multicellular organismen met vele celtypes, zodat is de controle van genuitdrukking veel ingewikkelder. Niet verrassend, wordt de genuitdrukking in eukaryotic cellen gecontroleerd door een aantal complexe processen die door de volgende lijst worden samengevat.
- na de bevruchting worden de cellen in het zich ontwikkelende embryo steeds specialer, voornamelijk door bepaalde genen aan te zetten en vele andere uit te schakelen., Sommige cellen in de alvleesklier, bijvoorbeeld, zijn gespecialiseerd om spijsverteringsenzymen te synthetiseren en af te scheiden, terwijl andere alvleeskliercellen (β-cellen in de eilandjes van Langerhans) gespecialiseerd zijn om insuline te synthetiseren en af te scheiden. Elk type van cel heeft een bepaald patroon van uitgedrukte genen. Deze differentiatie
 in gespecialiseerde cellen gebeurt grotendeels als gevolg van het uitschakelen van de expressie van de meeste genen in de cel; volwassen cellen kunnen slechts 3-5% van de genen in de celkern gebruiken.,
in gespecialiseerde cellen gebeurt grotendeels als gevolg van het uitschakelen van de expressie van de meeste genen in de cel; volwassen cellen kunnen slechts 3-5% van de genen in de celkern gebruiken., - genexpressie in eukaryoten kan ook worden gereguleerd door veranderingen in de verpakking van DNA, die de toegang van de transcriptie-enzymen van de cel (bijvoorbeeld RNA-polymerase) tot DNA moduleert. De illustratie hieronder toont aan dat chromosomen een complexe structuur hebben. De DNA-helix is gewikkeld rond speciale eiwitten die histonen worden genoemd, en dit is verpakt in strakke spiraalvormige vezels. Deze vezels worden vervolgens gelusd en gevouwen in steeds compactere structuren, die, wanneer volledig opgerold en gecondenseerd, de chromosomen hun karakteristieke verschijning geven in metafase.,
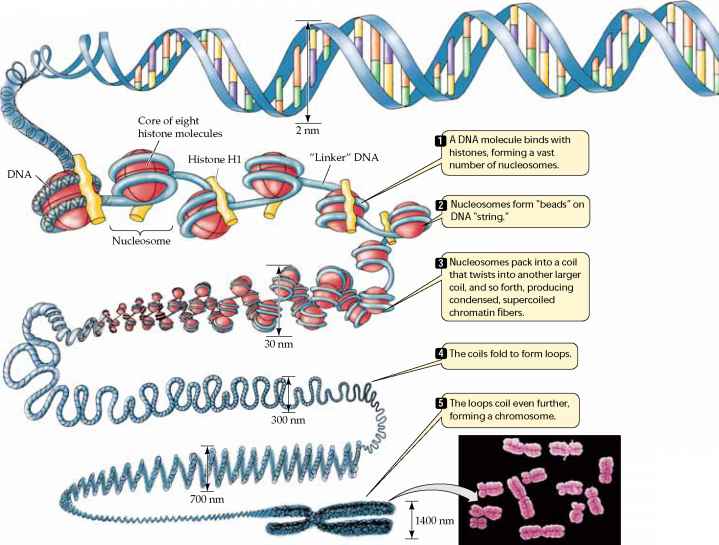
bron: http://www.78stepshealth.us/plasma-membrane/eukaryotic-chromosomes.html
- vergelijkbaar met de hierboven beschreven operons voor prokaryotes, eukaryotes ook gebruik regelgevende proteã nen om transcriptie te controleren, maar elk eukaryotic gen heeft zijn eigen reeks controles. Bovendien zijn er veel meer regelgevende proteã nen in eukaryotes en de interactie zijn veel complexer.,
- in eukaryotes vindt de transcriptie plaats binnen de membraangebonden kern, en het initiële transcript wordt gewijzigd voordat het van de kern naar het cytoplasma wordt getransporteerd voor vertaling in het ribosoom. het initiële transcript in eukaryotes heeft coderende segmenten (exons) afgewisseld met niet-coderende segmenten (introns)., Voordat het mRNA de kern verlaat, worden de introns uit het transcript verwijderd door een proces dat RNA-splicing wordt genoemd (zie onderstaande afbeelding & video), en worden extra nucleotiden toegevoegd aan de uiteinden van het transcript; deze niet-coderende “caps” en “staarten” beschermen het mRNA tegen aanvallen door cellulaire enzymen en helpen bij herkenning door de ribosomen.,
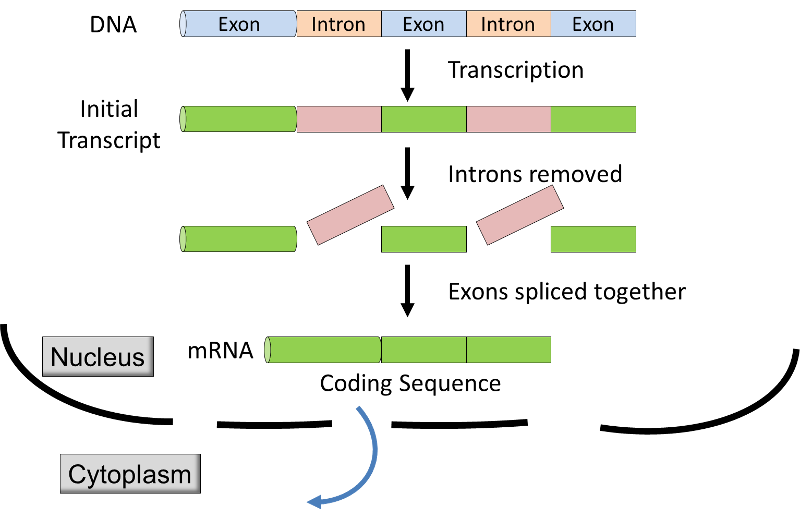
Bron: http://unmug.com/category/biology/organisation-control-of-genome/
![]()
- Variatie in de levensduur van mRNA biedt nog een andere mogelijkheid voor controle van de expressie van genen. Prokaryotic mRNA is zeer kortstondig, maar eukaryotic afschriften kunnen uren, of soms zelfs weken (b. v., mRNA voor hemoglobine in de rode bloedcellen van vogels) duren.,
- het vertaalproces biedt extra mogelijkheden voor Regulatie door veel eiwitten. Bijvoorbeeld, wordt de vertaling van hemoglobine mRNA geremd tenzij ijzer-bevattende heme in de cel aanwezig is.
- er zijn ook mogelijkheden voor “post-translationele” controles van genexpressie in eukaryoten. Sommige vertaalde polypeptiden (eiwitten) worden door enzymen in kleinere, actieve eindproducten gesneden. zoals geïllustreerd in de figuur hieronder die post-translationele verwerking van het hormoon insuline toont., Insuline wordt aanvankelijk vertaald als een grote, inactieve precursor; een signaalopeenvolging wordt verwijderd uit het hoofd van de precursor, en een groot centraal gedeelte (de C-keten) wordt weg gesneden, verlaten twee kleinere peptide kettingen die dan met elkaar door disulfidebruggen worden verbonden.De kleinere uiteindelijke vorm is de actieve vorm van insuline.
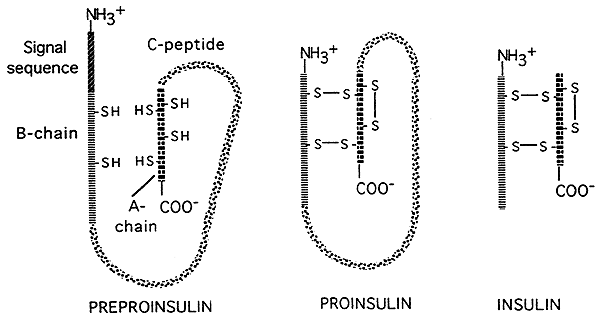
bron: http://www.nbs.csudh.edu/chemistry/faculty/nsturm/CHE450/19_InsulinGlucagon.htm
- genexpressie kan ook worden gewijzigd door de afbraak van de geproduceerde eiwitten., Bijvoorbeeld, worden sommige enzymen betrokken bij celmetabolisme afgebroken kort nadat zij worden geproduceerd; dit verstrekt een mechanisme om snel aan veranderende metabolische eisen te reageren.
- genexpressie kan ook worden beïnvloed door signalen van andere cellen. Er zijn vele voorbeelden waarin een signaalmolecuul (b.v., een hormoon) van één cel aan een receptorproteã NE op een doelcel bindt en een opeenvolging van biochemische veranderingen (een weg van de signaaltransductie) in werking stelt die in veranderingen binnen de doelcel resulteren., Deze veranderingen kunnen verhoogde of verminderde transcriptie omvatten zoals in onderstaande figuur wordt geïllustreerd.
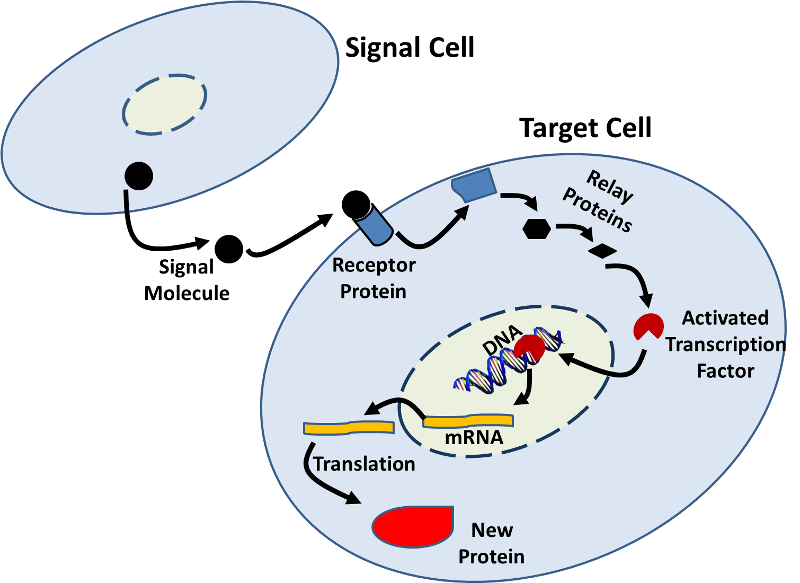
- het RNA-Interferentiesysteem (RNAi) is nog een ander mechanisme waarmee cellen de genexpressie controleren door de vertaling van mRNA af te sluiten. RNAi kan ook worden gebruikt om vertaling van virale proteã nen te sluiten wanneer een cel door een virus wordt besmet. Het RNAi-systeem heeft ook het potentieel om therapeutisch te worden geëxploiteerd.,
RNAi
sommige RNA-virussen zullen cellen binnendringen en dubbelstrengs RNA introduceren dat gebruik zal maken van de celmachines om nieuwe kopieën van viraal RNA en virale eiwitten te maken. Het RNA-interferentiesysteem van de cel (RNAi) kan het virale RNA verhinderen zich te herhalen. Eerst, hakt een enzym bijgenaamd “Dicer” om het even welk dubbelstrengs RNA het in stukken vindt die ongeveer 22 nucleotiden lang zijn. Vervolgens binden eiwitcomplexen RISC (RNA-induced silencing Complex) aan de fragmenten van double-stranded RNA, winden het, en geeft dan één van de bundels vrij, terwijl het behouden van de andere., Het RISC-RNA complex zal dan binden aan een ander viraal RNA met nucleotide sequenties die overeenkomen met die op het RNA in bijlage bij het complex. Deze binding blokkeert de vertaling van virale eiwitten ten minste gedeeltelijk, zo niet volledig. Het RNAi-systeem zou potentieel kunnen worden gebruikt om behandelingen voor gebrekkige genen te ontwikkelen die ziekte veroorzaken. De behandeling zou inhouden het maken van een dubbelstrengs RNA van het zieke gen en het introduceren van het in cellen om de uitdrukking van dat gen tot zwijgen te brengen., Voor een geïllustreerde uitleg van RNAi, zie de korte, interactieve Flash module op http://www.pbs.org/wgbh/nova/body/rnai-explained.html
het RNA interferentie systeem wordt ook vollediger uitgelegd in de video hieronder Van Nature Video.
![]()
![]()
return to top / previous page / next page