Ved genekspression menes transkriptionen af et gen til mRNA og dets efterfølgende oversættelse til protein. Genekspression styres primært på transkriptionsniveau, stort set som et resultat af binding af proteiner til specifikke steder på DNA. I 1965 Francois Jacob, Jacques Monod, og André Lwoff delte Nobelprisen i medicin for deres arbejde, der understøtter tanken om, at kontrol af enzymet i celler, der er reguleret af transkription af DNA., opstår gennem regulering af transkription, som enten kan induceres eller undertrykkes. Disse forskere foreslog, at produktionen af en .ymet styres af en “operon”, som består af en række beslægtede gener på kromosomet bestående af en operatør, en promotor, et regulatorgen og strukturelle gener.de strukturelle gener indeholder koden for de proteinprodukter, der skal produceres. Regulering af proteinproduktion opnås i vid udstrækning ved at modulere adgang af RNA-polymerase til det strukturelle gen, der transkriberes.,promotorgenet koder ikke for noget; det er simpelthen en DNA-sekvens, der er indledende bindingssted for RNA-polymerase.
- Eksempel på Inducible Transskription: bakterien E. coli, som har tre gener, der koder for enzymer, der gør det muligt at opdele og nedbryde lactose (sukker i mælk). Promotoren er stedet på DNA, hvor RNA-polymerase binder for at initiere transkription. Imidlertid er en .ymerne normalt til stede i meget lave koncentrationer, fordi deres transkription hæmmes af et represorprotein produceret af et regulatorgen (se den øverste del af figuren nedenfor)., Repressorproteinet binder til operatørstedet og hæmmer transkription. Men hvis lactose er til stede i miljøet, kan det binde til repressorproteinet og inaktivere det, effektivt at fjerne blokaden og muliggøre transkription af messenger RNA nødvendig til syntese af disse gener (nedre del af figuren nedenfor).

- Eksempel på Repressible Transskription: E. coli har brug for aminosyren tryptofan, og DNA ‘ et i E. coli også har gener til at syntetisere det., Disse gener transkriberer generelt kontinuerligt, da bakterien har brug for tryptophan. Hvis tryptophan-koncentrationerne imidlertid er høje, undertrykkes (slukkes) transkription ved at binde til et repressorprotein og aktivere det som illustreret nedenfor.
![]()
Kontrol af genekspression i Eukaryoter
Eukaryote celler, har lignende mekanismer til kontrol af genekspression, men de er mere komplekse., Overvej, for eksempel, at prokaryote celler af en bestemt art, er det samme, men de fleste eukaryoter er flercellede organismer med mange celletyper, så kontrol af genekspression er meget mere kompliceret. Ikke overraskende styres genekspression i eukaryote celler af en række komplekse processer, som opsummeres af følgende liste.
- efter befrugtning bliver cellerne i det udviklende embryo mere og mere specialiserede, stort set ved at tænde for nogle gener og slukke for mange andre., Nogle celler i bugspytkirtlen er for eksempel specialiserede til at syntetisere og udskille fordøjelsesen .ymer, mens andre bugspytkirtelceller (β-celler i Langerhans-øerne) er specialiserede til syntese og udskiller insulin. Hver type celle har et bestemt mønster af udtrykte gener. Denne differentiering
 i specialiserede celler forekommer stort set som et resultat af at slukke ekspressionen af de fleste gener i cellen; modne celler må kun bruge 3-5% af de gener, der er til stede i cellens kerne.,
i specialiserede celler forekommer stort set som et resultat af at slukke ekspressionen af de fleste gener i cellen; modne celler må kun bruge 3-5% af de gener, der er til stede i cellens kerne., - genekspression i eukaryoter kan også reguleres gennem ændringer i pakning af DNA, som modulerer adgang til cellens transskription enzymer (fx, RNA polymerase) til DNA. Illustrationen nedenfor viser, at kromosomer har en kompleks struktur. DNA-Heli .en er indpakket omkring specielle proteiner kaldet histoner, og dette er indpakket i stramme spiralfibre. Disse fibre Løkkes derefter og foldes ind i stadig mere kompakte strukturer, som, når de er fuldt opviklet og kondenseret, giver kromosomerne deres karakteristiske udseende i metafase.,
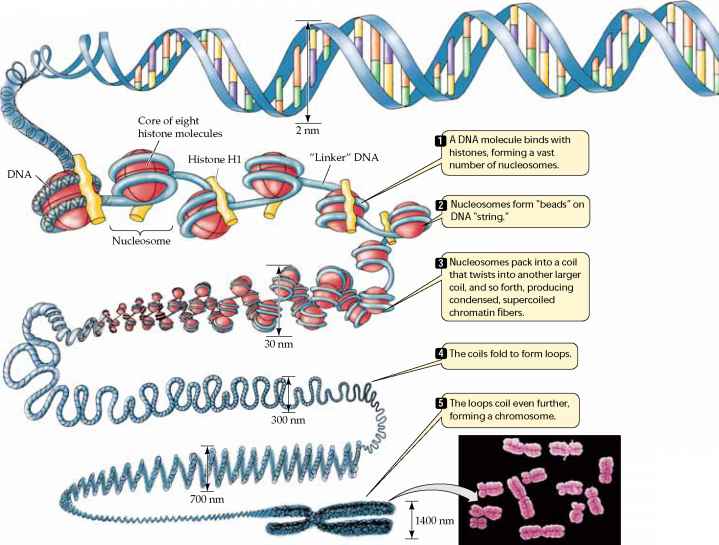
Kilde: http://www.78stepshealth.us/plasma-membrane/eukaryotic-chromosomes.html
- Svarer til operons beskrevet ovenfor for prokaryotes, eukaryoter også bruge regulatoriske proteiner til at styre transskription, men hver eukaryote gen har sin egen sæt af kontroller. Derudover er der mange flere regulatoriske proteiner i eukaryoter, og interaktionerne er meget mere komplekse.,
- I eukaryoter transskription finder sted inden for de membran-bundet kerne, og den oprindelige udskrift er ændret inden det transporteres fra kernen til cytoplasmaet for oversættelse ribosomets s. Den oprindelige udskrift i eukaryoter har kodning segmenter (exons) vekslende med ikke-kodende segmenter (introns)., Inden mRNA forlader cellekernen, introns, der er fjernet fra en afskrift af en proces kaldet RNA-splejsning (se grafisk & video nedenfor), og ekstra nukleotider, der er føjet til enderne af afskrift; disse ikke-kodende “caps” og “haler” beskytte mRNA fra angreb af cellulære enzymer og støtte i anerkendelse af ribosomer.,
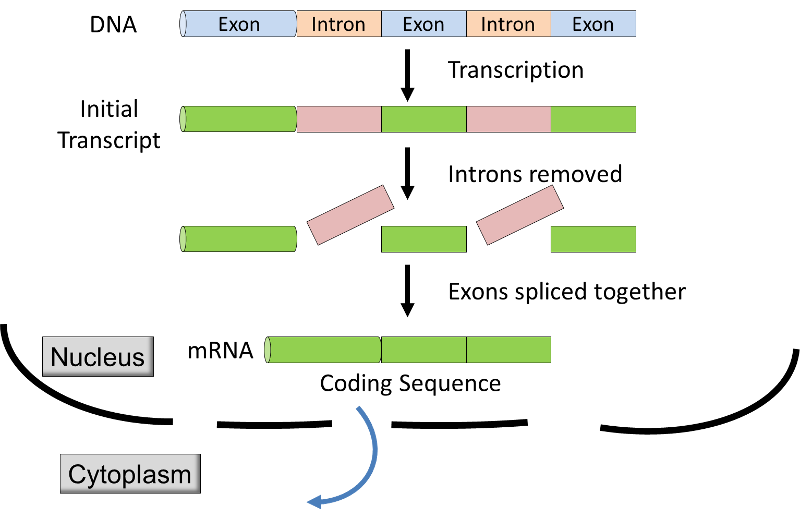
Kilde: http://unmug.com/category/biology/organisation-control-of-genome/
![]()
- Variation i levetiden af mRNA giver endnu en mulighed for kontrol af genekspression. Prokaryotisk mRNA er meget kortvarig, men eukaryote udskrifter kan vare timer, eller nogle gange endda uger (f.mRNA for hæmoglobin i de røde blodlegemer af fugle).,
- oversættelsesprocessen giver yderligere muligheder for regulering af mange proteiner. For eksempel hæmmes oversættelsen af hæmoglobin-mRNA, medmindre jernholdig hæm er til stede i cellen.
- Der er også muligheder for “post-translationelle” kontroller af genekspression i eukaryoter. Nogle oversatte polypeptider (proteiner) skæres af en .ymer i mindre aktive slutprodukter. som illustreret i figuren nedenfor, der viser post-translationel behandling af hormoninsulinet., Insulin oversættes oprindeligt som en stor, inaktiv forløber; en signalsekvens fjernes fra hovedet på forløberen, og en stor central del (C-kæden) skæres væk, hvilket efterlader to mindre peptidkæder, som derefter er forbundet med hinanden ved disulfidbroer.Den mindre endelige form er den aktive form af insulin.
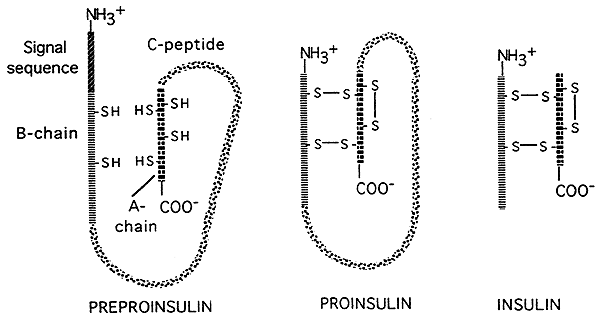
Kilde: http://www.nbs.csudh.edu/chemistry/faculty/nsturm/CHE450/19_InsulinGlucagon.htm
- genekspression kan også ændres ved fordelingen af de proteiner, der er produceret., For eksempel, nogle af de en .ymer, der er involveret i cellemetabolisme, nedbrydes kort efter, at de er produceret; dette giver en mekanisme til hurtigt at reagere på skiftende metaboliske krav.
- genekspression kan også påvirkes af signaler fra andre celler. Der er mange eksempler, hvor et signalmolekyle (f.et hormon) fra en celle binder til et receptorprotein på en målcelle og initierer en sekvens af biokemiske ændringer (en signaltransduktionsvej), der resulterer i ændringer i målcellen., Disse ændringer kan omfatte øget eller nedsat transskription som illustreret i figuren nedenfor.
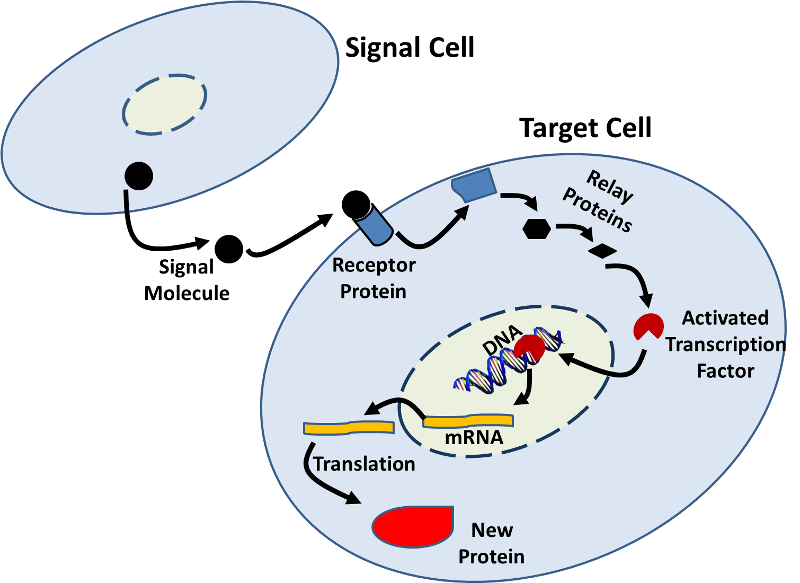
- RNA-Interferens system (rna interference (rnai)) er endnu en anden mekanisme, som celler kontrol af genekspression ved at lukke oversættelse af mRNA. RNAi kan også bruges til at lukke translation af virale proteiner, når en celle er inficeret af en virus. RNAi-systemet har også potentialet til at blive udnyttet terapeutisk.,
RNAi
nogle RNA-virus vil invadere celler og indføre dobbeltstrenget RNA, som vil bruge cellemaskineriet til at lave nye kopier af viralt RNA og virale proteiner. Cellens RNA-interferenssystem (RNAi) kan forhindre det virale RNA i at replikere. For det første hugger et en .ym med tilnavnet “Dicer” ethvert dobbeltstrenget RNA, det finder i stykker, der er omkring 22 nukleotider lange. Dernæst binder proteinkomplekser kaldet RISC (RNA-induceret Lyddæmpningskompleks) til fragmenterne af dobbeltstrenget RNA, vinder det og frigiver derefter en af strengene, samtidig med at den anden bevares., RISC-RNA-komplekset vil derefter binde til ethvert andet viralt RNA med nukleotidsekvenser, der matcher dem på RNA, der er knyttet til komplekset. Denne binding blokerer translation af virale proteiner i det mindste delvist, hvis ikke fuldstændigt. RNAi-systemet kan potentielt bruges til at udvikle behandlinger for defekte gener, der forårsager sygdom. Behandlingen ville involvere fremstilling af et dobbeltstrenget RNA fra det syge gen og introduktion af det i celler for at dæmpe ekspressionen af dette gen., For en illustreret forklaring af RNAi, se det korte, interaktive Flashmodul på http://www.pbs.org/wgbh/nova/body/rnai-explained.html
RNA-interferenssystemet forklares også mere fuldstændigt i videoen nedenfor fra Nature Video.
![]()
![]()
tilbage til top | forrige side | næste side