遺伝子発現によって、遺伝子のmRNAへの転写およびそれに続くタンパク質への翻訳を意味する。 遺伝子発現は、主に転写のレベルで制御され、主にDNA上の特定の部位へのタンパク質の結合の結果として制御される。 1965年、Francois Jacob、Jacques Monod、Andre Lwoffは、細胞内の酵素レベルの制御はDNAの転写によって調節されるという考えを支持する研究によってノーベル医学賞を受賞した。, 誘導または抑制することができる転写の調節を介して発生します。 これらの研究者らは、この酵素の産生は、オペレーター、プロモーター、調節遺伝子、および構造遺伝子からなる染色体上の一連の関連遺伝子からなる”オペロン”によって制御されることを提案した。
- 構造遺伝子は、産生されるタンパク質産物のコードを含む。 タンパク質産生の調節は、転写される構造遺伝子へのRNAポリメラーゼのアクセスを調節することによって主に達成される。,
- プロモーター遺伝子は何もコードしません。
- プロモーター遺伝子はRNAポリメラーゼの初期結合部位であるDNA配列に過ぎません。
- オペレーター遺伝子も非コーディングであり、それはリプレッサーの結合部位である単なるDNA配列である。
- レギュレーター遺伝子は、オペレーターに結合し、RNAポリメラーゼが構造遺伝子を転写するのをブロックするリプレッサー分子の合成をコードします。オペレーター遺伝子は、リプレッサー結合部位である非転写可能DNAの配列である。, また、hatがオペレーターに結合するリプレッサー分子の合成をコードするレギュレーター遺伝子もあります
- 誘導性転写の例:細菌E.coliは、乳糖(乳中の糖)を分割して代謝することを可能にする酵素をコードする三つの遺伝子を持っています。 プロモーターは、RNAポリメラーゼが転写を開始するために結合するDNA上の部位である。 しかし、これらの酵素は通常、レギュレータ遺伝子によって産生されるリプレッサータンパク質によって転写が阻害されるため、非常に低い濃度で存在する(下の図の上部を参照)。, リプレッサータンパク質はオペレーターサイトに結合し、転写を阻害する。 しかし、ラクトースが環境中に存在する場合、リプレッサータンパク質に結合して不活性化することができ、ブロックを効果的に除去し、これらの遺伝子の合成に必要なメッセンジャー RNAの転写を可能にする(下図の下の部分)。

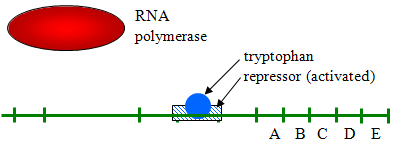
- 抑制性転写の例:大腸菌にはアミノ酸トリプトファンが必要であり、大腸菌のDNAにもそれを合成する遺伝子があります。, これらの遺伝子は、細菌がトリプトファンを必要とするので、一般に連続的に転写する。 しかし、トリプトファン濃度が高い場合、以下に示すように、リプレッサータンパク質に結合して活性化することによって、転写が抑制される(オフになる)。
真核生物における遺伝子発現の制御
真核細胞は、遺伝子発現の制御のための同様のメカニズムを有するが、それらはより複雑である。, 例えば、与えられた種の原核細胞はすべて同じであるが、ほとんどの真核生物は多くの細胞型を有する多細胞生物であるため、遺伝子発現の制御ははるかに複雑であると考えてください。 驚くことではないが、真核細胞における遺伝子発現は、以下のリストによって要約される多数の複雑なプロセスによって制御される。
- 受精後、発達中の胚の細胞は、主にいくつかの遺伝子をオンにし、他の多くの遺伝子をオフにすることによって、ますます特殊化されるようになる。, 例えば、膵臓の一部の細胞は消化酵素の合成と分泌に特化しており、他の膵細胞(ランゲルハンス島のβ細胞)はインスリンの合成と分泌に特化してい 各タイプの細胞は、発現された遺伝子の特定のパターンを有する。
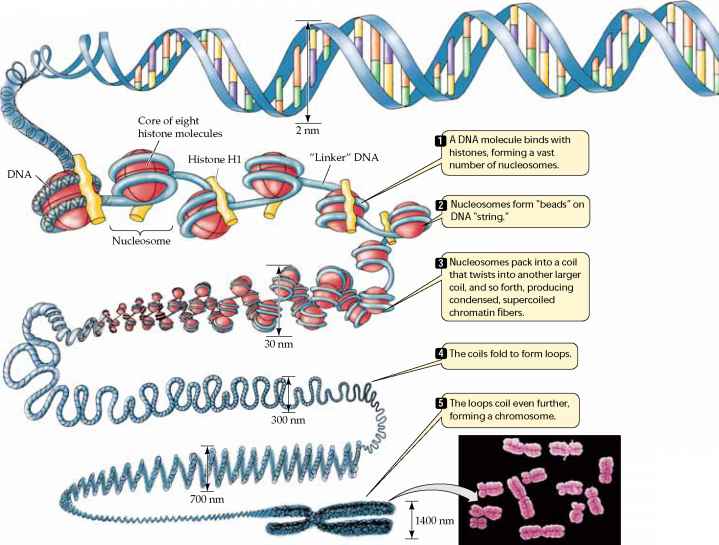
 特殊化された細胞へのこの分化は、主に細胞内のほとんどの遺伝子の発現をオフにした結果として起こり、成熟細胞は細胞の核に存在する遺伝子の3-5%しか使用できない。,真核生物における遺伝子発現はまた、細胞の転写酵素(例えば、RNAポリメラーゼ)のDNAへのアクセスを調節するDNAのパッキングの変化によって調節され 下の図は、染色体が複雑な構造を有することを示している。 DNAの螺旋はヒストンと呼ばれる特別な蛋白質のまわりで包まれ、これは堅い螺旋形繊維に包まれます。 その後、これらの繊維はループされ、ますますコンパクトな構造に折り畳まれ、完全にコイル状に凝縮されると、染色体に中期に特徴的な外観を与える。,
特殊化された細胞へのこの分化は、主に細胞内のほとんどの遺伝子の発現をオフにした結果として起こり、成熟細胞は細胞の核に存在する遺伝子の3-5%しか使用できない。,真核生物における遺伝子発現はまた、細胞の転写酵素(例えば、RNAポリメラーゼ)のDNAへのアクセスを調節するDNAのパッキングの変化によって調節され 下の図は、染色体が複雑な構造を有することを示している。 DNAの螺旋はヒストンと呼ばれる特別な蛋白質のまわりで包まれ、これは堅い螺旋形繊維に包まれます。 その後、これらの繊維はループされ、ますますコンパクトな構造に折り畳まれ、完全にコイル状に凝縮されると、染色体に中期に特徴的な外観を与える。,
ソース:http://www.78stepshealth.us/plasma-membrane/eukaryotic-chromosomes.html
- 原核生物については、上記のオペロンと同様に、真核生物も転写を制御するために調節タンパク質を使用するが、それぞれの真核生物遺伝子遺伝子は、転写を制御するために調節タンパク質を使用する。独自のコントロールセットがあります。 さらに、真核生物にはより多くの調節タンパク質があり、相互作用ははるかに複雑である。,
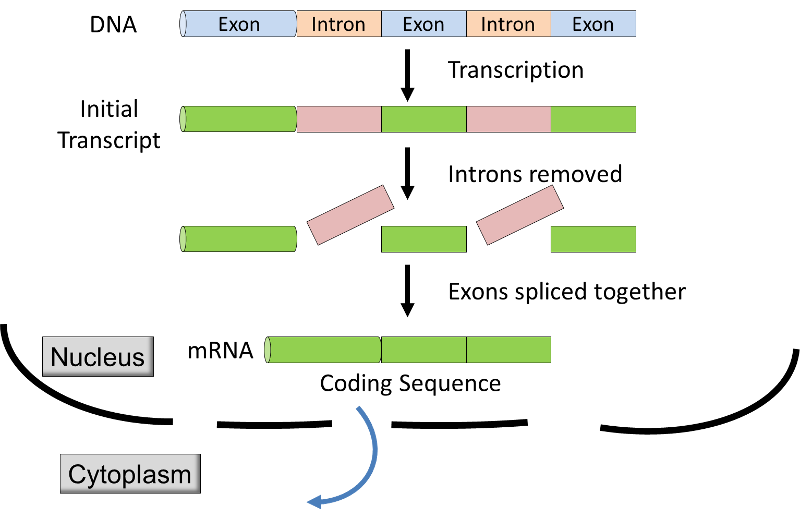
- 真核生物における転写は、膜に結合した核内で行われ、最初の転写産物は、リボソームsで翻訳のために核から細胞質に輸送される前に修飾される。, MRNAが核を離れる前に、RNAスプライシングと呼ばれるプロセスによってイントロンが転写産物から除去され(下の図&ビデオを参照)、余分なヌクレオチドが転写産物の端に追加される。,
ソース:http://unmug.com/category/biology/organisation-control-of-genome/
![]()
- mrnaの寿命の変化は、遺伝子発現の制御のためのさらに別の機会を提供します。 原核生物のmRNAは非常に短命であるが、真核生物の転写産物は数時間、または時には数週間持続することがある(例えば、鳥類の赤血球中のヘモグロビン,
- 翻訳のプロセスは、多くのタンパク質による調節のための追加の機会を提供する。 例えば、鉄containing有ヘムが細胞内に存在しない限り、ヘモグロビンmrnaの翻訳は阻害される。
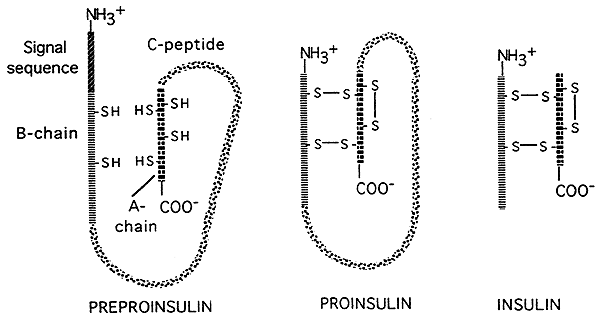
- 真核生物における遺伝子発現の”翻訳後”制御の機会もある。 いくつかの翻訳ポリペプチド(タンパク質)は、酵素によってより小さく活性な最終製品に切断される。 ホルモンのインシュリンの翻訳後の処理を描写する下の図で示されるように。, インスリンは最初に大きく不活性な前駆体として翻訳され、前駆体の頭部からシグナル配列が除去され、大きな中央部(C鎖)が切断され、二つの小さなペプチド鎖が残され、ジスルフィド橋によって互いに連結される。より小さい最終的な形態はインシュリンの活動的な形態です。
ソース:http://www.nbs.csudh.edu/chemistry/faculty/nsturm/CHE450/19_InsulinGlucagon.htm
- 遺伝子発現は、産生されるタンパク質の分解によって変更することもできる。, 例えば、細胞の新陳代謝にかかわる酵素のいくつかは作り出された直後に破壊されます;これは急速に変更の新陳代謝の要求に答えるためにメカ
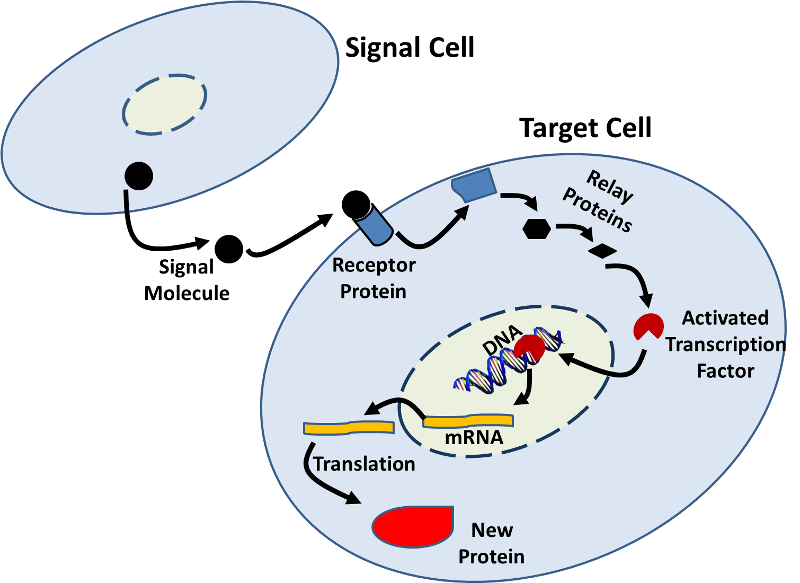
- 遺伝子発現は、他の細胞からのシグナルによって影響を受けることもできる。 ある細胞からのシグナル分子(例えば、ホルモン)が標的細胞上の受容体タンパク質に結合し、標的細胞内で変化をもたらす生化学的変化(シグナル伝達経路)のシーケンスを開始する例は多くある。, これらの変化には、下の図に示すように転写の増加または減少が含まれ得る。
- RNA干渉システム(RNAi)は、mRNAの翻訳を遮断することによって細胞が遺伝子発現を制御するさらに別のメカニズムである。 RNAiも使用可能シャットダウ翻訳のウイルスのタンパク質が、細胞がウィルスに感染する. Rnaiシステムはまた、治療的に利用される可能性を有する。,
RNAi
いくつかのRNAウイルスは、細胞に侵入し、ウイルスRNAおよびウイルスタンパク質の新しいコピーを作るために細胞機械を使用する二本鎖RNAを導入 細胞のRNA干渉システム(Rnai)は、ウイルスRNAの複製を防止することができる。 まず、”ダイサー”という愛称の酵素は、それが長さ約22ヌクレオチドである断片に見つけた任意の二本鎖RNAをチョップします。 次に、RISC(RNA誘導サイレンシング複合体)と呼ばれるタンパク質複合体は、二本鎖RNAの断片に結合し、巻き取り、他方を保持しながら一方の鎖を放出する。, その後、RISC-RNA複合体は、その複合体に結合したRNA上のヌクレオチド配列と一致する任意の他のウイルスRNAに結合する。 この結合は、ウイルスタンパク質の翻訳を少なくとも部分的にブロックします。 RNAiシステムは、疾患を引き起こす欠損遺伝子の治療法を開発するために潜在的に使用することができる。 治療は、疾患遺伝子から二本鎖RNAを作製し、その遺伝子の発現を抑制するためにそれを細胞に導入することを含むであろう。, RNAiの説明については、http://www.pbs.org/wgbh/nova/body/rnai-explained.html
RNA干渉システムについても、Nature Videoの下のビデオでより完全に説明されています。
![]()
![]()
トップに戻る|前のページ/次のページ