1. – Bildung
2. Struktur
3. Funktion
Mikrovilli sind kurze dünne Vorsprünge, die normalerweise in den freien Oberflächen einiger differenzierter Zellen vorkommen und häufiger in Epithelzellen vorkommen (Abbildungen 1 und 2). Sie sind filiforme Strukturen von etwa 1 bis 2 µm Länge und etwa 100 nm Dicke. Im Inneren enthält jede Mikrovillosis Dutzende von Aktinfilamenten, die parallel zur Langachse ausgerichtet sind., Mikrovilli sind normalerweise dicht und dicht gepackt wie ein Bürstenrand. In der Draufsicht sind Mikrovilli räumlich als Sechsecke organisiert.

Abbildung 1. Dünndarmepithel bei Lichtmikroskopie (Bild links) und Rasterelektronenmikroskopie (Bild rechts) mit Mikrovilli an der freien Oberfläche der Zellen.
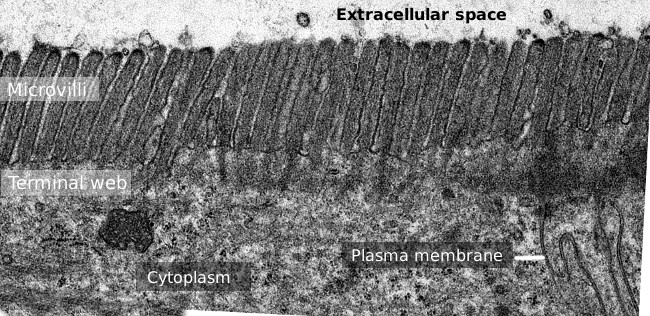
Abbildung 2. Transmissionselektronenmikroskopie Bild der Dünndarmepitheloberfläche., Das terminale Netz von Aktinfilamenten ist das dunklere Band, das basal zu Mikrovilli beobachtet wird.
Es gibt viele Zelltypen, die Mikrovilli tragen, aber sie werden häufiger in Epithelzellen gefunden, wie Darm-Enterozyten, gewundenes Gangepithel von Nieren und Nebenhoden. Mikrovilli können auch in einigen spezialisierten sensorischen Zellen wie Riechrezeptorneuronen, Organen von Cortis Rezeptorzellen, Plazentazellen und in sich bewegenden Zellen gefunden werden., Obwohl Mikrovilli in verschiedenen Zelltypen eine ähnliche äußere Morphologie aufweisen, enthalten sie eine unterschiedliche molekulare Zusammensetzung und zeigen einige Unterschiede in ihrer inneren Struktur. Im Folgenden beschäftigen wir uns mit den Merkmalen von Mikrovilli von Enterozyten des Dünndarmepithels.
Bildung
Mikrovilli werden gebildet, nachdem Aktinfilamente mit einer dichten Platte in der zytosolischen Oberfläche der Plasmamembran assoziiert sind. Proteine zum Sammeln von Aktinfilamenten in Bündeln und deren Stabilisierung werden ebenfalls benötigt. Es sieht so aus, als wäre es ein sequentieller Prozess., Erstens erleichtert Villin die Bildung von Bündeln von Aktinfilamenten und stabilisiert ihre distalen Enden, die das Filamentwachstum hemmen, so dass Villin wichtig sein kann, um die Länge von Mikrovilli zu bestimmen. Ezrin, weniger reichlich vorhanden, hilft, die Aktinfilamente des Bündels mit der Plasmamembran zu verbinden. Eine zweite Phase der Mikrovilli-Bildung umfasst das Erreichen der endgültigen Länge und die Verteilung auf der Zelloberfläche als Sechsecke. Fimbrin und Spin können zu diesen Merkmalen beitragen. Laterale Verbindungen durch Protocadherine zwischen Aktinfilamenten verschiedener Mikrovilli treten auch während dieser zweiten Stufe auf., Es ist wichtig zu beachten, dass während der Mikrovillienbildung eine große Menge an Membran-und Membranproteinen benötigt wird, die durch eine intensive Exozytose zugeführt wird. Evaginationen der Plasmamembran, eine pro Mikrovillosität, ist eine Folge der Aktin Filamentpolymerisation.
Struktur
Mikrovilli bestehen aus 6 Hauptproteinen: Aktin, Fimbrin, Villin, Myosin (Myo1A), Calmodulin und Spectrin (nicht erythrozytisch) (Abbildung 3). Das Mikrovellositäts-Skelett ist ein Bündel von 30-40 Aktin-Filamenten parallel zur langen Achse und mit ihren Minusenden zur Spitze der Mikrovellosität hin., Fimbrin und Villin verbinden die Aktinfilamente miteinander, während Myo1A und Calmodulin die Verbindungen zwischen dem Bündel und der Membran herstellen. Das Plus-Ende der Aktinfilamente erstrecken sich in das Zytoplasma und dort vermischen sich die Aktinfilamente verschiedener Mikrovilli untereinander und bilden ein Netz mit einem hexagonalen Muster. Dieses Netz wird als terminales Netz bezeichnet und breitet sich durch die Zellperipherie unter den Mikrovilli aus. Terminal Web enthält eine große Menge an nicht-erythrozytischem Spectrin.
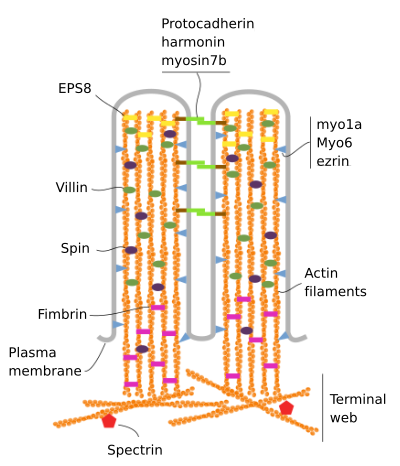
Abbildung 3., Molekulare Organisation von Mikrovilli (angepasst von Crawley et al., 2014)
Jede Mikrovellosität ist stabil und bewegt sich nicht, aber ihr Proteinskelett wird permanent erneuert, indem Aktin und andere Gerüstproteine zwischen der Mikrovellosität und dem Cytosol ausgetauscht werden. Es bedeutet, dass es ein Gleichgewicht zwischen eingehenden und ausgehenden Proteinen gibt. Es wird geschätzt, dass alle 20 Minuten das Skelett jeder Mikrovellosität vollständig erneuert wird. Mikrovilli können vollständig unorganisiert sein und nach einer Erhöhung der Kalziumkonzentration verschwinden, beispielsweise unter Zellstress., Calcium verändert das Verhalten von Villin, von stabilisierenden zu trennenden Aktinfilamenten. Mikrovilli verschwinden in Zellen, die in die Mitose eintreten. Die Terminalbahn ist auch eine Kunststoffstruktur.
Funktion
Der Austausch von Substanzen zwischen Hohlräumen oder äußeren Umgebungen und Geweben ist eine der Hauptfunktionen von Epithelien wie Darm-und Nierenepithelien. Es wird hauptsächlich an der apikalen (freien) Oberfläche der Epithelzellen durchgeführt, wo sich die meisten molekularen Transporter, Ionenpumpen und endozytären Prozesse befinden., Je mehr Zelloberfläche, desto mehr Platz für die molekularen Maschinen für den Austausch. Mikrovilli sind wirbelsäulenartige Vorsprünge, die die Plasmamembranoberfläche und damit den Membranproteingehalt enorm erhöhen. Dies ist besonders wichtig bei jenen absorptiven und sekretorischen Zellen von Epithelien. Mikrovilli können die Zelloberfläche im Vergleich zu einer flachen Membran um das 100-fache vergrößern. Enterozyten-Mikrovilli tragen viele Enzyme, die nicht am molekularen Austausch, sondern an der Verdauung beteiligt sind.
Mikrovilli können molekulare Transduktionswege beeinflussen., Ihre Membranen arbeiten als relativ isolierte Domänen mit einem unterschiedlichen Satz von Molekülen, wie Glukosetransportern, Ionenpumpen und Rezeptoren. Die Länge der Mikrovilli ist gut geeignet, um die lokale Transduktion der molekularen Bahnen unabhängig vom Zytosol durchzuführen. Darüber hinaus bilden die Aktinfilamente und Strukturproteine ein dichtes Netzwerk von Molekülen, die irgendwie die Moleküle filtern oder steuern, die das Innere jeder Mikrovellosität kommen oder verlassen. Dieses Gerüst kann auch als zeitlicher Speicher für Kalzium dienen.,
Die hohe Dichte von Mikrovilli bildet eine physikalische Barriere, die vor Krankheitserregern schützt. Dies ist wichtig für das Darmepithel. Darüber hinaus ist die große Menge an Membran, die in Mikrovilli gespeichert ist, ein Reservoir, das hypertonischen Beleidigungen standhält und den Bruch der Plasmamembran verhindert.
Stereozilien sind spezialisierte Mikrovillositäten, die sensorische Funktionen ausführen. Trotz des Namens stereocilium sind sie tatsächlich modifizierte Mikrovillositäten, die in sensorische Strukturen umgewandelt werden, und einige Autoren schlagen stattdessen den Namen estereomicrovellosity vor., Sie können im Nebenhodenepithel und im Innenohr gefunden werden und arbeiten als Mechanorezeptoren, die die umgebende Flüssigkeitsbewegung spüren. Sterozilien des Säugetier-Innenohrs finden sich als Teil des Corti-Organs. Sie sind 10 bis 50 µm lang und enthalten mehr als 3000 Aktinfilamente im Inneren. Die Schallwellen der Luft werden in Flüssigkeitswellen im Ohr umgewandelt, die Stereozilien wahrnehmen und in elektrische Signale umwandeln, die über den Hörnerv zum Gehirn gelangen. Stereocilia sind wie Antennen, die externe Signale empfangen, die in elektrischen Informationen transduziert werden.,
Einige Mikrovellositäten sind auf das Erfassen von Licht spezialisiert. Photorezeptoren sind lichtsensorische Zellen, die in Sehorganen wie der Netzhaut vorkommen. Die zellulären sensorischen Strukturen sind tatsächlich modifizierte Zilien oder Mikrovellositäten. Photorezeptoren mit modifizierten Mikrovellositäten zur Lichterkennung finden sich in visuellen Strukturen von Wirbellosen. Diese Photorezeptoren enthalten Mikrovellositäten, die in den sogenannten Rhabdomeren angeordnet sind. Jede Mikrovellosität enthält visuelle Pigmente, die auf niedrige Lichtintensitäten reagieren können und bei hohen Lichtintensitäten effizient sind., Mikrovellositäten, die das sogenannte Rhabdomer bilden, und der molekulare Transduktionsweg machen die mikrovillositätsbasierte Lichtreaktion sinnvoller als Cilium-lichtsensorische Systeme von Wirbeltieren.
Es wird vorgeschlagen, eine große Anzahl extrazellulärer Vesikel aus Mikrovilli freizusetzen. Die Spitzen der Enterozytenmikrovilli sind in der Lage, Vesikel in die Umgebung freizusetzen., Die Verbindung zwischen Plasmamembran und Aktin-Myosin-Netzwerk kann dazu beitragen, diese Vesikel zu produzieren, indem die Membran in Richtung des distalen Teils jeder Mikrovellosität gezogen wird, wo kleine Teile der Membran in Vesikel umgewandelt werden. Diese extrazellulären Vesikel enthalten Enzyme und sind mit alkalischer Phosphatase angereichert.
Literaturverzeichnis
Brown J W, McKnight, C J. 2010. Molekulares Modell des mikrovillären Zytoskeletts und Organisation der Bürstengrenze. PLoS One. 5: e940 ![]() .
.
Crawley SW, Mooseker MS, Tyska MJ. 2014. Die Gestaltung der intestinalen bürstensaum., J Zelle Biol. 207: 441-451 ![]() .
.
Fain G L, Hardie R, Laughlin S, B. 2010. Phototransduktion und die Entwicklung von Photorezeptoren. Curr Biol. 20: R114-R124 ![]() .
.
Lange, K. 2011. Grundlegende Rolle von Mikrovilli in den Hauptfunktionen differenzierter Zellen: Umriss eines universellen Regulierungs-und Signalsystems an der Zellperipherie. J Zelle Physiol. 226: 896-92
, McConnell R E, Higginbotham J N, Shifrin Jr, D A, Tabb D L, Coffey, R J, Tyska M J. 2009. Der Enterozyten-Mikrovillus ist eine Vesikel erzeugende Organelle. J Zelle Biol., 185: 1285-129 ![]() .
.
Sauvanet C, Wayt J, Pelaseyed T, Bretscher, A. 2015. Struktur, Regulation und funktionelle Vielfalt von Mikrovilli auf der apikalen Domäne von Epithelzellen. Annual review of cell and development biology. 31: 593-621.