1. Vorming
2. Structuur
3. Functie
Microvilli zijn korte dunne uitsteeksels die gewoonlijk worden gevonden in de vrije oppervlakken van sommige gedifferentieerde cellen, frequenter in epitheelcellen (figuren 1 en 2). Het zijn filiforme structuren van ongeveer 1 tot 2 µm lang en ongeveer 100 nm dik. Binnen, bevat elke microvillosity tientallen actin filamenten die parallel aan de lange as worden georiënteerd., Microvilli zijn meestal dicht en strak verpakt als een borstelrand. In bovenaanzicht zijn microvilli ruimtelijk georganiseerd als zeshoeken.

figuur 1. Klein intestinaal epitheel bij lichtmicroscopie (afbeelding links) en bij scanning elektronenmicroscopie (afbeelding rechts) met microvilli aan het vrije oppervlak van de cellen.
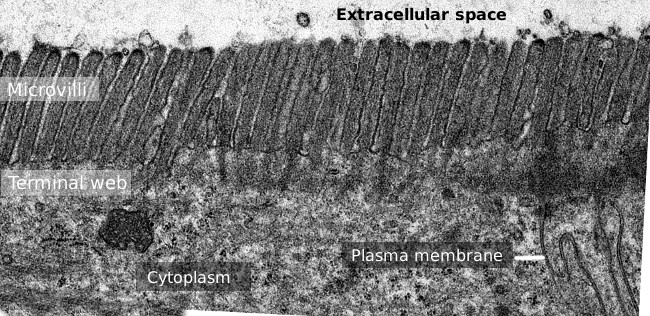
Figuur 2. Transmissie elektronenmicroscopie beeld van de dunne darm epitheel oppervlak., Het eind web van actine filamenten is de donkerdere band waargenomen basaal aan microvilli.
Er zijn veel celtypen die microvilli bevatten, maar ze worden vaker aangetroffen in epitheelcellen, zoals darm enterocyten, ingewikkeld ductepitheel van nieren en epididymus. Microvilli kan ook worden gevonden in sommige gespecialiseerde sensorische cellen zoals olfactorische receptor neuronen, orgaan van Corti receptor cellen, placenta cellen, en in cellen die bewegen., Hoewel microvilli vergelijkbare externe morfologie in verschillende celtypes tonen, bevatten zij verschillende moleculaire samenstelling en tonen sommige verschillen in hun interne structuur. Hieronder hebben we te maken met de kenmerken van microvilli van enterocyten van het epitheel van de dunne darm.
vorming
Microvillen worden gevormd nadat actine filamenten worden geassocieerd met een dichte plaat die wordt aangetroffen in het cytosolisch oppervlak van het plasmamembraan. De proteã nen voor het verzamelen van actin filamenten in bundels en het stabiliseren van hen zijn ook nodig. Het lijkt erop dat het een sequentieel proces is., Ten eerste, villin vergemakkelijkt de vorming van bundels actine filamenten en stabiliseert hun distale uiteinden remmen de gloeidraad groeien, zodat villin kan belangrijk zijn om de lengte van microvilli vast te stellen. Ezrin, minder overvloedig, helpt om de actin gloeidraden van de bundel aan het plasmamembraan te verbinden. Een tweede fase van microvilli-vorming omvat het bereiken van de uiteindelijke lengte en de distributie op de celoppervlakte als zeshoeken. Fimbrin en spin kunnen bijdragen aan deze functies. Zijverbindingen door protocadherins tussen actin gloeidraden van verschillende microvilli gebeuren ook tijdens dit tweede stadium., Het is belangrijk op te merken dat tijdens de vorming van microvilli een grote hoeveelheid membraan-en membraaneiwitten nodig is, die wordt geleverd door een intense exocytose. Evaginations van plasmamembraan, één per elke microvillosity, is een gevolg van actin filament polymerisatie.
structuur
Microvilli bestaan uit 6 belangrijke eiwitten: actine, fimbrine, villin, myosine (Myo1A), calmoduline en spectrin (niet-erythrocytisch) (Figuur 3). Het microvellosity skelet is een bundel van 30-40 actine filamenten evenwijdig aan de lange as en met hun min uiteinden in de richting van de punt van de microvellosity., Fimbrine en villin verbinden samen de actine filamenten, terwijl Myo1A en calmodulin de verbindingen tussen de bundel en het membraan maken. Het plus eind van actin gloeidraden strekt zich uit in het cytoplasma en daar mengen de actin gloeidraden van verschillende microvilli zich tussen elkaar en vormen een web met een hexagonaal patroon. Dit web staat bekend als terminaal web, en het verspreidt zich door de celrand onder de microvilli. Terminaal web bevat grote hoeveelheid niet-erytrocytaire spectrin.
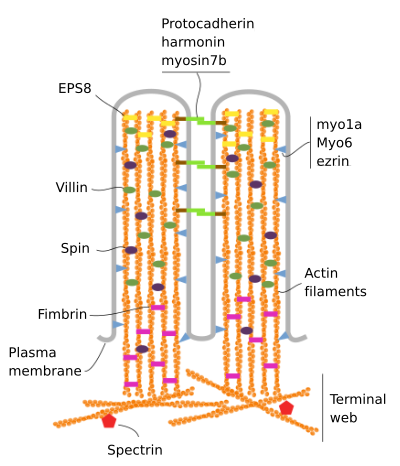
Figuur 3., Moleculaire organisatie van microvilli (aangepast van Crawley et al., 2014)
elke microvellositeit is stabiel en beweegt niet, maar zijn proteïneskelet is in permanente vernieuwing door het uitwisselen van actine-en andere steigereiwitten tussen de microvellositeit en het cytosol. Het betekent dat er een evenwicht is tussen inkomende en uitgaande eiwitten. Geschat wordt dat elke 20 minuten het skelet van elke microvellositeit wordt vernieuwd. Microvilli kan volledig ongeorganiseerd zijn en verdwijnen na een verhoging van de calciumconcentratie, bijvoorbeeld onder celstress., Calcium verandert het gedrag van villin, van stabiliserend tot het scheiden van actine filamenten. Microvilli verdwijnen in cellen die de mitose ingaan. Het terminalweb is ook een kunststof structuur.
functie
uitwisseling van stoffen tussen holtes of uitwendige omgeving en weefsels is een van de belangrijkste functies van epithelia, zoals darm-en nierepithelia. Het wordt meestal gedaan op het apicale (vrije) oppervlak van de epitheliale cellen, waar de meeste moleculaire transporter, ionenpompen zijn gevestigd, en endocytische processen gebeuren., Hoe meer celoppervlak hoe meer ruimte om de moleculaire machines voor de uitwisseling onder te brengen. Microvilli zijn stekelachtige uitsteeksels die het plasmamembraan oppervlak en daarmee het membraaneiwitgehalte enorm verhogen. Dit bijzonder belangrijk in die absorberende en secretorische cellen van epithelia. Microvilli kan het celoppervlak tot 100 keer verhogen in vergelijking met een vlak membraan. Enterocyt microvilli dragen veel enzymen, die niet betrokken zijn bij moleculaire uitwisseling, maar bij de spijsvertering.
Microvilli kunnen moleculaire transductieroutes beïnvloeden., Hun membranen werken als relatief geïsoleerde domeinen met een aparte reeks moleculen, zoals glucosetransporters, ionenpompen en receptoren. De lengte van microvilli is goed geschikt voor het uitvoeren van lokale transductie van de moleculaire routes onafhankelijk van cytosol. Bovendien vormen de actin gloeidraden en de structurele proteã NEN steiger een dicht netwerk van molecules dat op de een of andere manier filtert of de molecules controleert die komen of het verlaten van het binnenland van elke microvellosity. Deze steiger kan ook werken als tijdelijke opslag voor calcium.,
De hoge dichtheid van microvilli vormt een fysieke barrière die beschermt tegen pathogenen. Dit is belangrijk voor darmepitheel. Bovendien is de grote hoeveelheid membraan opgeslagen in microvilli een reservoir dat hypertone beledigingen kan weerstaan en de breuk van het plasmamembraan kan voorkomen.
Stereocilia zijn gespecialiseerde microvilloben die zintuiglijke functies vervullen. Ondanks de naam stereocilium, ze zijn eigenlijk gemodificeerde microvillosities omgezet in zintuiglijke structuren,en sommige auteurs stellen de naam estereomicrovellosity in plaats daarvan., Ze kunnen worden gevonden in de epitheel epitheel, en in het binnenoor, werken als mechanoreceptoren die de omliggende vloeibare beweging voelen. Sterocilia van het binnenoor van het zoogdier worden gevonden Als onderdeel van het Corti-orgaan. Ze zijn 10 tot 50 µm lang en bevatten meer dan 3000 actine filament binnenin. De geluidsgolven van de lucht worden getransformeerd in vloeibare golven in het oor, die stereocilia voelen en omzetten in elektrische signalen die via de gehoorzenuw naar de hersenen reizen. Stereocilia zijn als antennes die externe signalen ontvangen, die worden omgezet in elektrische informatie.,
sommige microvellen zijn gespecialiseerd in het detecteren van licht. Fotoreceptoren zijn licht sensorische cellen gevonden in visuele organen zoals het netvlies. De cellulaire sensorische structuren zijn eigenlijk gemodificeerde trilharen of microvellosities. Fotoreceptoren met gemodificeerde microvellen voor lichtdetectie kunnen worden gevonden in ongewervelde visuele structuren. Deze fotoreceptoren bevatten microvellen gerangschikt in de zogenaamde rhabdomeren. Elke microvellositeit bevat visuele pigmenten die kunnen reageren op lage lichtintensiteit en efficiënt zijn bij hoge lichtintensiteit., Microvellosities organiseren de vorming van de zogenaamde rhabdomere, en de transductie moleculaire route maken microvillosity gebaseerde lichtrespons verstandiger dan cilium licht sensorische systemen van gewervelde dieren.
een groot aantal extracellulaire blaasjes zou uit microvilli vrijkomen. De uiteinden van enterocyt microvilli kunnen blaasjes aan het omringende milieu vrijgeven., De verbinding tussen plasmamembraan en actin-myosin netwerk kan helpen om deze blaasjes te produceren door membraan naar het distale deel van elke microvellosity te slepen, waar de kleine delen van membraan in blaasjes worden getransformeerd. Deze extracellulaire blaasjes bevatten enzymen en zijn verrijkt met alkalische fosfatase.
Bibliography
Brown J W, McKnight C J. 2010. Moleculair model van het microvillar cytoskelet en organisatie van de borstelrand. PLoS ÉÉN. 5: e940 ![]() . Crawley SW, Mooseker MS, Tyska MJ. 2014. Vormgeven van de darm borstel grens., J Cell Biol. 207: 441-451
. Crawley SW, Mooseker MS, Tyska MJ. 2014. Vormgeven van de darm borstel grens., J Cell Biol. 207: 441-451 ![]() .
.
Fain G L, Hardie R, Laughlin s B. 2010. Fototransductie en de evolutie van fotoreceptoren. Curr Biol. 20: R114-R124 ![]() .
.
Lange K. 2011. Fundamentele rol van microvilli in de belangrijkste functies van gedifferentieerde cellen: overzicht van een universeel regelend en signalerend systeem aan de celrand. J Cell Physiol. 226: 896-92
McConnell R E, Higginbotham J N, Shifrin Jr D A, Tabb D L, Coffey R J, Tyska M J. 2009. Enterocyte microvillus is een vesikel-genererende organelle. J Cell Biol., 185: 1285-129 ![]() .
.
Sauvanet C, Wayt J, Pelaseyed T, Bretscher A. 2015. Structuur, regelgeving en functionele diversiteit van microvilli op het apicale domein van epitheliale cellen. Jaarlijkse evaluatie van cel-en ontwikkelingsbiologie. 31: 593-621.