1. Formazione
2. Struttura
3. Funzione
I microvilli sono brevi sporgenze sottili che si trovano solitamente nelle superfici libere di alcune cellule differenziate, più frequenti nelle cellule epiteliali (Figure 1 e 2). Sono strutture filiformi di circa 1-2 µm di lunghezza e circa 100 nm di spessore. All’interno, ogni microvillosità contiene dozzine di filamenti di actina orientati parallelamente all’asse lungo., I microvilli sono solitamente densi e ben imballati come un bordo a pennello. Nella vista dall’alto, i microvilli sono organizzati spazialmente come esagoni.

Figura 1. Piccolo epitelio intestinale alla microscopia ottica (immagine a sinistra) e alla microscopia elettronica a scansione (immagine a destra) che mostra microvilli sulla superficie libera delle cellule.
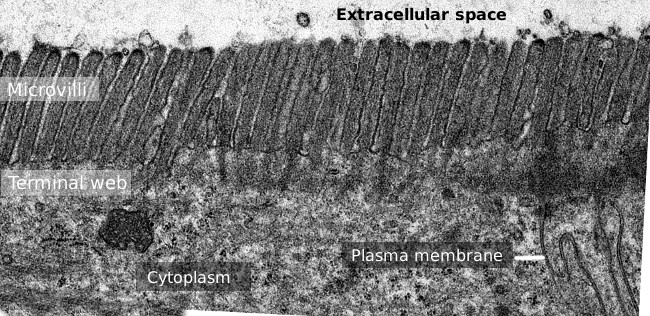
Figura 2. Microscopia elettronica a trasmissione immagine della superficie dell’epitelio dell’intestino tenue., La rete terminale dei filamenti di actina è la banda più scura osservata basale ai microvilli.
Ci sono molti tipi di cellule che portano microvilli, ma si trovano più frequentemente nelle cellule epiteliali, come gli enterociti intestinali, l’epitelio del dotto contorto dei reni e l’epididimo. I microvilli possono anche essere trovati in alcune cellule sensoriali specializzate come i neuroni del recettore olfattivo, l’organo delle cellule del recettore di Corti, le cellule della placenta e nelle cellule che si muovono., Sebbene i microvilli mostrino una morfologia esterna simile in diversi tipi di cellule, contengono una composizione molecolare distinta e mostrano alcune differenze nella loro struttura interna. Di seguito, abbiamo a che fare con le caratteristiche dei microvilli degli enterociti dell’epitelio dell’intestino tenue.
Formazione
I microvilli si formano dopo che i filamenti di actina si associano a una piastra densa trovata nella superficie citosolica della membrana plasmatica. Sono necessarie anche proteine per la raccolta di filamenti di actina in fasci e la loro stabilizzazione. Sembra che sia un processo sequenziale., In primo luogo, villin facilita la formazione di fasci di filamenti di actina e stabilizza le loro estremità distali inibendo la crescita del filamento, quindi villin può essere importante per stabilire la lunghezza dei microvilli. Ezrin, meno abbondante, aiuta a collegare i filamenti di actina del fascio alla membrana plasmatica. Una seconda fase della formazione dei microvilli comprende il raggiungimento della lunghezza finale e la distribuzione sulla superficie cellulare come esagoni. Fimbrin e spin possono contribuire a queste caratteristiche. Connessioni laterali da protocadherine tra filamenti di actina di diversi microvilli avvengono anche durante questo secondo stadio., È importante notare che durante la formazione dei microvilli è necessaria una grande quantità di proteine di membrana e di membrana, che viene fornita da un’intensa esocitosi. Le evaginazioni della membrana plasmatica, una per ogni microvillosità, sono una conseguenza della polimerizzazione del filamento di actina.
Struttura
I microvilli sono costituiti da 6 proteine principali: actina, fimbrina, villina, miosina (Myo1A), calmodulina e spettrina (non eritrocitaria) (Figura 3). Lo scheletro di microvellosità è un fascio di 30-40 filamenti di actina paralleli all’asse lungo e con le loro estremità negative verso la punta della microvellosità., Fimbrin e villin collegano insieme i filamenti di actina, mentre Myo1A e calmodulin fanno i collegamenti tra il fascio e la membrana. L’estremità più dei filamenti di actina si estende nel citoplasma e lì i filamenti di actina di diversi microvilli si mescolano tra loro e formano una rete con un motivo esagonale. Questa rete è nota come rete terminale e si diffonde attraverso la periferia della cellula sotto i microvilli. Il web terminale contiene la grande quantità di spectrin non erythrocytic.
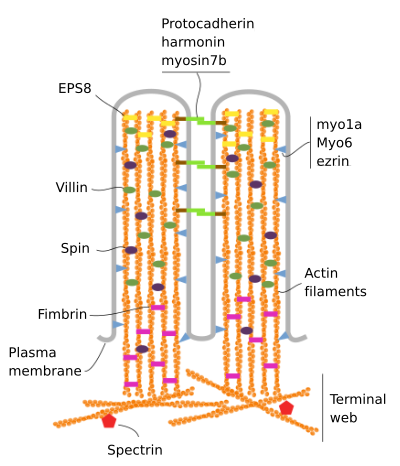
Figura 3., Organizzazione molecolare dei microvilli (adattato da Crawley et al., 2014)
Ogni microvellosità è stabile e non si muove, ma il suo scheletro proteico è in rinnovamento permanente scambiando actina e altre proteine di scaffold tra la microvellosità e il citosol. Significa che esiste un equilibrio tra proteine in entrata e in uscita. Si stima che ogni 20 minuti lo scheletro di ogni microvellosità sia completato rinnovato. I microvilli possono essere completamente disorganizzati e scomparire dopo un aumento della concentrazione di calcio, ad esempio sotto stress cellulare., Il calcio cambia il comportamento della villina, dalla stabilizzazione alla rottura dei filamenti di actina. I microvilli scompaiono nelle cellule che entrano nella mitosi. Il web terminale è anche una struttura in plastica.
Funzione
Lo scambio di sostanze tra cavità o ambiente esterno e tessuti è una delle principali funzioni degli epiteli, come gli epiteli intestinali e renali. È per lo più fatto sulla superficie apicale (libera) delle cellule epiteliali, dove si trovano la maggior parte dei trasportatori molecolari, le pompe ioniche e i processi endocitici., Più superficie cellulare più spazio per ospitare il macchinario molecolare per lo scambio. I microvilli sono sporgenze simili a spine che aumentano enormemente la superficie della membrana plasmatica e quindi il contenuto proteico della membrana. Questo è particolarmente importante in quelle cellule assorbenti e secretorie di epiteli. I microvilli possono aumentare la superficie cellulare fino a 100 volte rispetto a una membrana piatta. I microvilli degli enterociti portano molti enzimi, che non sono coinvolti nello scambio molecolare, ma nella digestione.
I microvilli possono influenzare le vie molecolari di trasduzione., Le loro membrane funzionano come domini relativamente isolati con un insieme distinto di molecole, come trasportatori di glucosio, pompe ioniche e recettori. La lunghezza dei microvilli è adatta per eseguire la trasduzione locale delle vie molecolari indipendentemente dal citosol. Inoltre, i filamenti di actina e l’impalcatura delle proteine strutturali formano una fitta rete di molecole che in qualche modo filtra o controlla le molecole che entrano o escono dall’interno di ogni microvellosità. Questa impalcatura può anche funzionare come deposito temporale per il calcio.,
L’alta densità dei microvilli costituisce una barriera fisica che protegge dagli agenti patogeni. Questo è importante per l’epitelio intestinale. Inoltre, la grande quantità di membrana immagazzinata nei microvilli è un serbatoio in grado di sopportare insulti ipertonici e prevenire la rottura della membrana plasmatica.
Le stereocilia sono microvillosità specializzate che svolgono funzioni sensoriali. Nonostante il nome stereocilium, sono in realtà microvillosità modificate trasformate in strutture sensoriali, e alcuni autori propongono invece il nome estereomicrovellosity., Possono essere trovati nell’epitelio dell’epididimo e nell’orecchio interno, lavorando come meccanorecettori che percepiscono il movimento fluido circostante. Sterocilia dell’orecchio interno del mammifero si trovano come parte dell’organo del Corti. Sono da 10 a 50 µm di lunghezza e contengono più di 3000 filamenti di actina all’interno. Le onde sonore dell’aria si trasformano in onde fluide nell’orecchio, che stereocilia senso e trasformare in segnali elettrici che viaggia attraverso il nervo uditivo al cervello. Le stereocilia sono come antenne che ricevono segnali esterni, che vengono trasdotti in informazioni elettriche.,
Alcune microvellosità sono specializzate nel rilevamento della luce. I fotorrecettori sono cellule sensoriali leggere che si trovano negli organi visivi come la retina. Le strutture sensoriali cellulari sono in realtà ciglia modificate o microvellosità. I fotorecettori con microvellosità modificate per il rilevamento della luce possono essere trovati nelle strutture visive degli invertebrati. Quei fotorecettori contengono microvellosità disposte nei cosiddetti rabdomeri. Ogni microvellosità contiene pigmenti visivi che possono rispondere a basse intensità di luce e sono efficienti ad alte intensità di luce., Le microvellosità si organizzano formando il cosiddetto rabdomo e la via molecolare di trasduzione rende la risposta alla luce basata sulla microvillosità più sensibile dei sistemi sensoriali leggeri del ciglio dei vertebrati.
Si propone di rilasciare un gran numero di vescicole extracellulari dai microvilli. Le punte dei microvilli enterocitari sono in grado di rilasciare vescicole nell’ambiente circostante., La connessione tra la membrana plasmatica e la rete di actina-miosina può aiutare a produrre queste vescicole trascinando la membrana verso la parte distale di ciascuna microvellosità, dove piccole porzioni di membrana vengono trasformate in vescicole. Queste vescicole extracellulari contengono enzimi e sono arricchite in fosfatasi alcalina.
Bibliografia
Brown J W, McKnight C J. 2010. Modello molecolare del citoscheletro microvillare e organizzazione del bordo pennello. PLoS Uno. 5: e940 ![]() . per maggiori informazioni:
. per maggiori informazioni: ![]() .
.
Fain G L, Hardie R, Laughlin S B. 2010. Fototrasduzione ed evoluzione dei fotorecettori. Curr Biol. 20: R114-R124 ![]() .
.
Lange K. 2011. Ruolo fondamentale dei microvilli nelle principali funzioni delle cellule differenziate: Cenni di un sistema universale di regolazione e segnalazione alla periferia cellulare. Physiol delle cellule di J. 226: 896-92
McConnell R E, Higginbotham J N, Shifrin Jr D A, Tabb D L, Coffey R J, Tyska M J. 2009. Il microvillo degli enterociti è un organello che genera vescicole. Biol delle cellule J., 185: 1285-129 ![]() .
.
Sauvanet C, Wayt J, Pelaseyed T, Bretscher A. 2015. Struttura, regolazione e diversità funzionale dei microvilli sul dominio apicale delle cellule epiteliali. Revisione annuale della biologia cellulare e dello sviluppo. 31: 593-621.