1. Formation
2. Structure
3. Function
les microvillosités sont des protubérances courtes et minces que l’on trouve généralement dans les surfaces libres de certaines cellules différenciées, plus fréquentes dans les cellules épithéliales (Figures 1 et 2). Ce sont des structures filiformes d’environ 1 à 2 µm de long et d’environ 100 nm d’épaisseur. À l’intérieur, chaque microvillosité contient des dizaines de filaments d’actine orientés parallèlement au grand axe., Les microvillosités sont généralement denses et bien tassées comme une bordure de brosse. En vue de dessus, les microvillosités sont organisées spatialement en hexagones.

la Figure 1. Petite épithélium intestinal en microscopie optique (image de gauche) et au microscope électronique à balayage (image de droite) montrant des microvillosités à la surface libre des cellules.
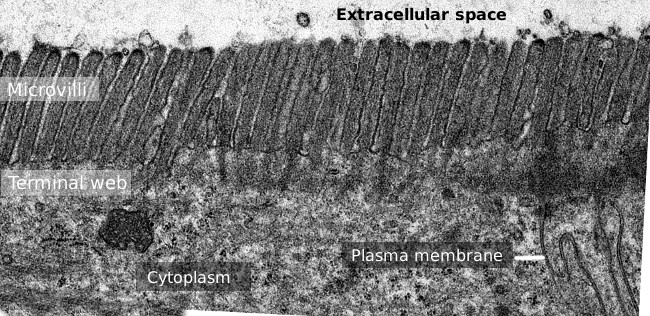
la Figure 2. Image de microscopie électronique à Transmission de la surface de l’épithélium de l’intestin grêle., La bande terminale des filaments d’actine est la bande la plus foncée observée de la base aux microvillosités.
Il existe de nombreux types de cellules portant des microvillosités, mais on les trouve plus fréquemment dans les cellules épithéliales, comme les entérocytes intestinaux, l’épithélium des conduits alambiqués des reins et l’épididyme. Les microvillosités peuvent également être trouvées dans certaines cellules sensorielles spécialisées comme les neurones récepteurs olfactifs, les cellules réceptrices de L’organe de Corti, les cellules du placenta et dans les cellules en mouvement., Bien que les microvillosités présentent une morphologie externe similaire dans différents types de cellules, elles contiennent une composition moléculaire distincte et présentent certaines différences dans leur structure interne. Ci-dessous, nous traitons des caractéristiques des microvillosités des entérocytes de l’épithélium de l’intestin grêle.
Formation
les microvillosités se forment après l’association de filaments d’actine à une plaque dense présente dans la surface cytosolique de la membrane plasmique. Des protéines pour rassembler les filaments d’actine en faisceaux et les stabiliser sont également nécessaires. On dirait que c’est un processus séquentiel., Tout d’abord, la villine facilite la formation de faisceaux de filaments d’actine et stabilise leurs extrémités distales en inhibant la croissance du filament, de sorte que la villine peut être importante pour établir la longueur des microvillosités. Ezrin, moins abondant, aide à connecter les filaments d’actine du faisceau à la membrane plasmique. Une deuxième phase de formation de microvillosités consiste à atteindre la longueur finale et la distribution à la surface de la cellule sous forme d’hexagones. Fimbrin et spin peuvent contribuer à ces caractéristiques. Les connexions latérales par les protocadhérines entre les filaments d’actine de différents microvillosités se produisent également au cours de cette deuxième étape., Il est important de noter que lors de la formation de microvillosités, une grande quantité de protéines membranaires et membranaires est nécessaire, qui est fournie par une exocytose intense. Les évaginations de la membrane plasmique, une par microvillosité, sont une conséquence de la polymérisation du filament d’actine.
Structure
les microvillosités sont constituées de 6 protéines principales: actine, fimbrine, villine, myosine (Myo1A), calmoduline et spectrine (Non érythrocytaire) (Figure 3). Le squelette de microvellosité est un faisceau de 30-40 filaments d’actine parallèles à l’axe long et avec leurs extrémités moins vers l’extrémité de la microvellosité., La fimbrine et la villine lient les filaments d’actine, tandis que la Myo1A et la calmoduline font les connexions entre le faisceau et la membrane. L’extrémité plus des filaments d’actine s’étend dans le cytoplasme et là, les filaments d’actine de différents microvillosités s’entremêlent entre eux et forment une bande avec un motif hexagonal. Ce web est connu sous le nom de web terminal, et il se propage à travers la périphérie cellulaire sous les microvillosités. Terminal web contient une grande quantité de spectrine Non érythrocytaire.
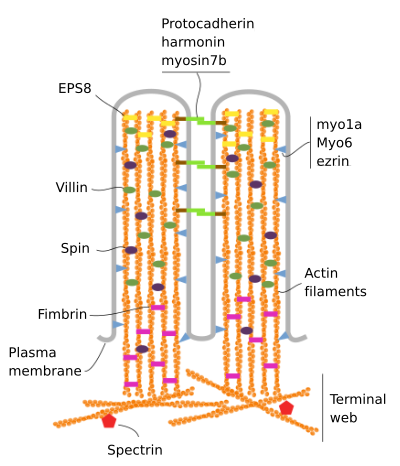
la Figure 3., Organisation moléculaire des microvillosités (adapté de Crawley et al., 2014)
chaque microvellosité est stable et ne bouge pas, mais son squelette protéique est en renouvellement permanent par échange d’actine et d’autres protéines d’échafaudage entre la microvellosité et le cytosol. Cela signifie qu’il existe un équilibre entre les protéines entrantes et sortantes. On estime que toutes les 20 minutes le squelette de chaque microvellosité est complété renouvelé. Les microvillosités peuvent être complètement désorganisées et disparaître après une augmentation de la concentration en calcium, par exemple sous stress cellulaire., Le Calcium modifie le comportement de la villine, de la stabilisation à la séparation des filaments d’actine. Les microvillosités disparaissent dans les cellules entrant dans la mitose. Le web terminal est également une structure en plastique.
fonction
L’échange de substances entre les cavités ou l’environnement externe et les tissus est l’une des principales fonctions des épithéliums, tels que les épithéliums intestinaux et rénaux. Il se fait principalement à la surface apicale (libre) des cellules épithéliales, où se trouvent la plupart des transporteurs moléculaires, des pompes ioniques et des processus endocytaires., Plus il y a de surface cellulaire, plus il y a d’espace pour accueillir la machinerie moléculaire pour l’échange. Les microvillosités sont des protubérances en forme de colonne vertébrale qui augmentent énormément la surface de la membrane plasmique et donc la teneur en protéines membranaires. Ceci est particulièrement important dans les cellules absorbantes et sécrétoires de l’épithélium. Les microvillosités peuvent augmenter la surface cellulaire jusqu’à 100 fois par rapport à une membrane plate. Les microvillosités entérocytaires portent de nombreuses enzymes, qui ne sont pas impliquées dans l’échange moléculaire, mais dans la digestion.
les microvillosités peuvent influencer les voies moléculaires de transduction., Leurs membranes fonctionnent comme des domaines relativement isolés avec un ensemble distinct de molécules, comme les transporteurs de glucose, les pompes ioniques et les récepteurs. La longueur des microvillosités est bien adaptée pour effectuer une transduction locale des voies moléculaires indépendamment du cytosol. De plus, les filaments d’actine et les protéines structurales forment un réseau dense de molécules qui filtrent ou contrôlent en quelque sorte les molécules venant ou quittant l’intérieur de chaque microvellosité. Cet échafaudage peut également fonctionner comme un stockage temporel pour le calcium.,
la haute densité de microvillosités constitue une barrière physique qui protège contre les agents pathogènes. Ceci est important pour l’épithélium intestinal. De plus, la grande quantité de membrane stockée dans les microvillosités est un réservoir qui peut résister aux insultes hypertoniques et empêcher la rupture de la membrane plasmique.
Les stéréocils sont des microvillosités spécialisées qui remplissent des fonctions sensorielles. Malgré le nom stereocilium, ce sont en fait des microvillosités modifiées transformées en structures sensorielles, et certains auteurs proposent plutôt le nom estereomicrovellosity., Ils peuvent être trouvés dans l’épithélium de l’épididyme et dans l’oreille interne, agissant comme des mécanorécepteurs qui détectent le mouvement du fluide environnant. Les stérociles de l’oreille interne des mammifères font partie de l’organe de Corti. Ils ont une longueur de 10 à 50 µm et contiennent plus de 3000 filaments d’actine à l’intérieur. Les ondes sonores de l’air sont transformées en ondes fluides dans l’oreille, que les stéréocils détectent et transforment en signaux électriques qui se déplacent via le nerf auditif vers le cerveau. Les stéréocils sont comme des antennes qui reçoivent des signaux externes, qui sont transductés en Informations électriques.,
certaines microvellosités sont spécialisées dans la détection de la lumière. Les photorécepteurs sont des cellules sensorielles de la lumière présentes dans les organes visuels comme la rétine. Les structures sensorielles cellulaires sont en fait des cils modifiés ou des microvellosités. Des photorécepteurs portant des microvellosités modifiées pour la détection de la lumière peuvent être trouvés dans les structures visuelles des invertébrés. Ces photorécepteurs contiennent des microvellosités disposées dans les soi-disant rhabdomères. Chaque microvellosité contient des pigments visuels qui peuvent répondre à de faibles intensités lumineuses et sont efficaces à des intensités lumineuses élevées., Les microvellosités s’organisent pour former le soi-disant rhabdomère, et la voie moléculaire de transduction rend la réponse à la lumière basée sur la microvillosité plus sensible que les systèmes sensoriels de la lumière du cil des vertébrés.
Il est proposé qu’un grand nombre de vésicules extracellulaires soient libérées à partir de microvillosités. Les extrémités des microvillosités entérocytaires sont capables de libérer des vésicules dans l’environnement environnant., La connexion entre la membrane plasmique et le réseau actine-myosine peut aider à produire ces vésicules en faisant glisser la membrane vers la partie distale de chaque microvellosité, où de petites portions de membrane sont transformées en vésicules. Ces vésicules extracellulaires contiennent des enzymes et sont enrichies en phosphatase alcaline.
Bibliographie
Marron J W, McKnight C J. 2010. Modèle moléculaire du cytosquelette microvillaire et organisation de la bordure en brosse. PLoS One. 5: e940 ![]() .
.
Crawley SW, MOOSEKER MS, Tyska MJ. 2014. Façonner la bordure de la brosse intestinale., J Cell Biol. 207: 441-451 ![]() .
.
Fain G L, Hardie R, Laughlin s B. 2010. Phototransduction et évolution des photorécepteurs. Curr Biol. 20: R114-R124 ![]() .
.
Lange K. 2011. Rôle fondamental des microvillosités dans les fonctions principales des cellules différenciées: Contour d’un système universel de régulation et de signalisation à la périphérie des cellules. J Cellule Physiol. 226: 896-92
McConnell R E, Higginbotham J N, Shifrin Jr D A, Tabb D L, Coffey R J, Tyska M J. 2009. Le microvillus entérocytaire est un organite produisant des vésicules. J Cell Biol., 185: 1285-129 ![]() .
.
Sauvanet C, Wayt J, Pelaseyed t, Bretscher A. 2015. Structure, régulation et diversité fonctionnelle des microvillosités sur le domaine apical des cellules épithéliales. Examen annuel de la cellule et de biologie du développement. 31: 593-621.