1. Formación
2. Estructura
3. Función
las microvellosidades son protuberancias cortas y delgadas que suelen encontrarse en las superficies libres de algunas células diferenciadas, más frecuentes en las células epiteliales (Figuras 1 y 2). Son estructuras filiformes de aproximadamente 1 a 2 µm de largo y alrededor de 100 nm de espesor. En el interior, cada microvilosidad contiene docenas de filamentos de actina orientados paralelos al eje largo., Las microvellosidades son generalmente densas y apretadas como un borde de pincel. En la vista superior, las microvellosidades están organizadas espacialmente como hexágonos.

la Figura 1. Epitelio del intestino delgado en microscopía óptica (imagen a la izquierda) y en microscopía electrónica de barrido (imagen a la derecha) mostrando microvellosidades en la superficie libre de las células.
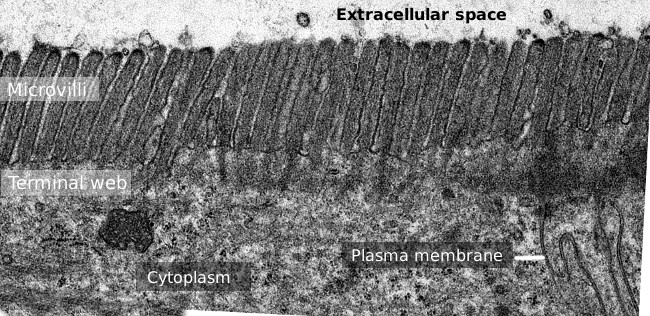
la Figura 2. Imagen de microscopía electrónica de transmisión de la superficie del epitelio del intestino delgado., La banda terminal de los filamentos de actina es la banda más oscura observada basal a microvellosidades.
hay muchos tipos de células que llevan microvellosidades, pero se encuentran con más frecuencia en las células epiteliales, como los enterocitos intestinales, el epitelio del conducto enrevesado de los riñones y el epidídimo. Las microvellosidades también se pueden encontrar en algunas células sensoriales especializadas como las neuronas receptoras olfativas, el órgano de las células receptoras de Corti, las células de la placenta y las células que se están moviendo., Aunque las microvellosidades muestran una morfología externa similar en diferentes tipos celulares, contienen una composición molecular distinta y muestran algunas diferencias en su estructura interna. A continuación, estamos tratando con las características de las microvellosidades de los enterocitos del epitelio del intestino delgado.
formación
las microvellosidades se forman después de que los filamentos de actina se asocian con una placa densa que se encuentra en la superficie citosólica de la membrana plasmática. También se necesitan proteínas para reunir los filamentos de actina en haces y estabilizarlos. Parece que es un proceso secuencial., En primer lugar, la vellina facilita la formación de haces de filamentos de actina y estabiliza sus extremos distales inhibiendo el crecimiento del filamento, por lo que la vellina puede ser importante para establecer la longitud de las microvellosidades. Ezrin, menos abundante, ayuda a conectar los filamentos de actina del haz a la membrana plasmática. Una segunda fase de la formación de microvellosidades incluye alcanzar la longitud final y la distribución en la superficie celular como hexágonos. Fimbrina y spin pueden contribuir a estas características. Las conexiones laterales por protocadherinas entre filamentos de actina de diferentes microvellosidades también ocurren durante esta segunda etapa., Es importante notar que durante la formación de microvellosidades se necesita una gran cantidad de membrana y proteínas de membrana, que es suministrada por una exocitosis intensa. Las evaginaciones de la membrana plasmática, una por cada microvilosidad, es una consecuencia de la polimerización del filamento de actina.
Estructura
las microvellosidades están compuestas por 6 proteínas principales: actina, fimbrina, vellina, miosina (Myo1A), calmodulina y espectrina (no eritrocítica) (Figura 3). El esqueleto de la microvelosidad es un haz de 30-40 filamentos de actina paralelos al eje largo y con sus extremos negativos hacia la punta de la microvelosidad., La fimbrina y la vellina unen los filamentos de actina, mientras que la Myo1A y la calmodulina hacen las conexiones entre el haz y la membrana. El extremo positivo de los filamentos de actina se extiende en el citoplasma y allí los filamentos de actina de diferentes microvellosidades se entremezclan entre sí y forman una red con un patrón hexagonal. Esta red se conoce como red terminal, y se propaga a través de la periferia celular bajo las microvellosidades. La banda Terminal contiene una gran cantidad de espectrina no eritrocítica.
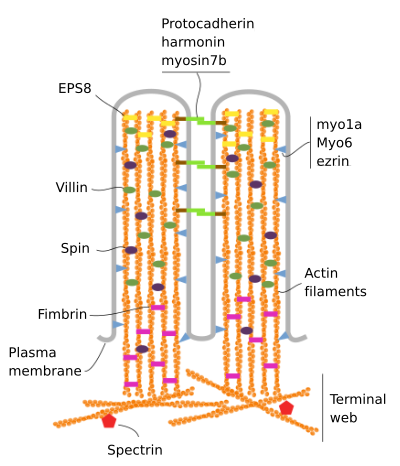
la Figura 3., Organización Molecular de microvellosidades (adaptado de Crawley et al., 2014)
cada microvelosidad es estable y no se mueve, pero su esqueleto proteico está en permanente renovación mediante el intercambio de actina y otras proteínas de andamio entre la microvelosidad y el citosol. Significa que hay un equilibrio entre las proteínas entrantes y salientes. Se estima que cada 20 minutos el esqueleto de cada microvellosidad se completa renovado. Las microvellosidades pueden estar completamente desorganizadas y desaparecer después de un aumento en la concentración de calcio, por ejemplo bajo estrés celular., El calcio cambia el comportamiento de la vellina, desde la estabilización hasta la ruptura de los filamentos de actina. Las microvellosidades desaparecen en las células que entran en la mitosis. La banda terminal es también una estructura de plástico.
función
El intercambio de sustancias entre las cavidades o el entorno externo y los tejidos es una de las principales funciones de los epitelios, como los epitelios de intestino y riñón. Se realiza principalmente en la superficie apical (libre) de las células epiteliales, donde se encuentran la mayoría de los transportadores moleculares, bombas de iones y ocurren procesos endocíticos., Cuanta más superficie celular, más espacio para acomodar la maquinaria molecular para el intercambio. Las microvellosidades son protuberancias en forma de columna vertebral que aumentan enormemente la superficie de la membrana plasmática y, por lo tanto, el contenido de proteínas de la membrana. Esto es particularmente importante en aquellas células absorbentes y secretoras de epitelios. Las microvellosidades pueden aumentar la superficie celular hasta 100 veces en comparación con una membrana plana. Las microvellosidades del enterocito tener muchas enzimas, que no están involucradas en el intercambio molecular, pero en la digestión.
las microvellosidades pueden influir en las vías moleculares de transducción., Sus membranas funcionan como dominios relativamente aislados con un conjunto distinto de moléculas, como transportadores de glucosa, bombas de iones y receptores. La longitud de las microvellosidades es muy adecuada para realizar la transducción local de las vías moleculares independientemente del citosol. Además, los filamentos de actina y las proteínas estructurales forman una densa red de moléculas que de alguna manera filtra o controla las moléculas que vienen o salen del interior de cada microvelosidad. Este andamio también puede funcionar como un almacenamiento temporal para el calcio.,
la alta densidad de microvellosidades hace una barrera física que protege contra patógenos. Esto es importante para el epitelio intestinal. Además, la gran cantidad de membrana almacenada en las microvellosidades es un reservorio que puede soportar insultos hipertónicos y evitar la rotura de la membrana plasmática.
Los Estereocilios son microvillosidades especializadas que realizan funciones sensoriales. A pesar del nombre stereocilium, en realidad son microvillosidades modificadas transformadas en estructuras sensoriales, y algunos autores proponen el nombre estereomicrovellosity en su lugar., Se pueden encontrar en el epitelio del epidídimo y en el oído interno, trabajando como mecanorreceptores que detectan el movimiento del fluido circundante. Los esterocilios del oído interno de los mamíferos se encuentran como parte del órgano del Corti. Tienen una longitud de 10 a 50 µm y contienen más de 3000 filamentos de actina en su interior. Las ondas sonoras del aire se transforman en ondas fluidas en el oído, que los estereocilios detectan y transforman en señales eléctricas que viajan a través del nervio auditivo al cerebro. Los estereocilios son como antenas que reciben señales externas, que se transducen en información eléctrica.,
algunas microvelosidades están especializadas en la detección de la luz. Los fotorreceptores son células sensoriales de luz que se encuentran en órganos visuales como la retina. Las estructuras sensoriales celulares son en realidad cilios modificados o microvelosidades. Los fotorreceptores con microvelosidades modificadas para la detección de luz se pueden encontrar en estructuras visuales de invertebrados. Esos fotorreceptores contienen microvelosidades dispuestas en los llamados rabdómeros. Cada microvelosidad contiene pigmentos visuales que pueden responder a intensidades de luz bajas y son eficientes a intensidades de luz altas., Las microvelosidades se organizan formando el llamado rabdómero, y la vía molecular de transducción hace que la respuesta a la luz basada en la microvilosidad sea más sensible que los sistemas sensoriales de luz cilium de los vertebrados.
se propone liberar un gran número de vesículas extracelulares de las microvellosidades. Las puntas de las microvellosidades de los enterocitos son capaces de liberar vesículas al entorno circundante., La conexión entre la membrana plasmática y la red actina-miosina puede ayudar a producir estas vesículas arrastrando la membrana hacia la parte distal de cada microvelosidad, donde pequeñas porciones de membrana se transforman en vesículas. Estas vesículas extracelulares contienen enzimas y están enriquecidas en fosfatasa alcalina.
Bibliografía
Brown J W, McKnight C J. 2010. Modelo Molecular del citoesqueleto microvilar y organización del borde del cepillo. PLoS UNO. 5: e940 ![]() .
.
Crawley SW, Mooseker MS, Tyska MJ. 2014. Dar forma al borde del cepillo intestinal., J Cell Biol. 207: 441-451 ![]() .
.
Fain G L, Hardie R, Laughlin s B. 2010. Fototransducción y evolución de los fotorreceptores. Curr Biol. 20: R114-R124 ![]() .
.
Lange K. 2011. Papel Fundamental de las microvellosidades en las principales funciones de las células diferenciadas: esquema de un sistema universal de regulación y señalización en la periferia celular. J Cell Physiol. 226: 896-92
McConnell R E, Higginbotham J N, Shifrin Jr D A, Tabb D L, Coffey R J, Tyska M J. 2009. Las microvellosidades del enterocito es una vesícula de generación de orgánulo. J Cell Biol., 185: 1285-129 ![]() .Sauvanet C, Wayt J, Pelaseyed T, Bretscher A. 2015. Estructura, regulación y diversidad funcional de las microvellosidades en el dominio apical de las células epiteliales. Annual review of cell and development biology. 31: 593-621.
.Sauvanet C, Wayt J, Pelaseyed T, Bretscher A. 2015. Estructura, regulación y diversidad funcional de las microvellosidades en el dominio apical de las células epiteliales. Annual review of cell and development biology. 31: 593-621.