1. Bildning
2. Struktur
3. Funktion
mikrovilli är korta tunna utsprång som vanligtvis finns i de fria ytorna hos vissa differentierade celler, oftare i epitelceller (figurerna 1 och 2). De är filiforma strukturer av ca 1 till 2 µm lång och runt 100 nm tjock. Inuti innehåller varje mikrovillositet dussintals aktinfilament orienterade parallellt med den långa axeln., Mikrovilli är vanligtvis täta och tätt packade som en borstgräns. I toppvy är mikrovilli rumsligt organiserade som hexagoner.

Figur 1. Tunntarmen epitel vid ljusmikroskopi (bild till vänster) och vid scanning elektronmikroskopi (bild till höger) visar mikrovilli på den fria ytan av cellerna.
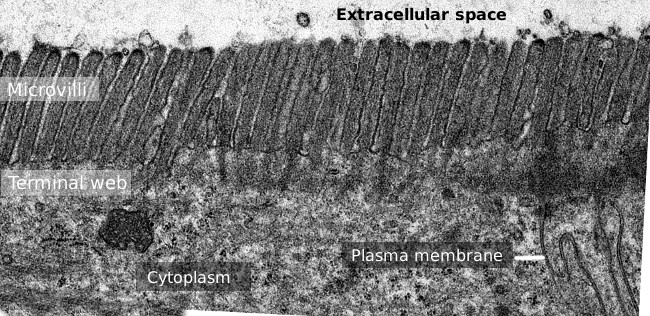
Figur 2. Transmissionselektronmikroskopi bild av tunntarmen epitelytan., Terminalbanan av aktinfilament är det mörkare bandet observerade basal till mikrovilli.
det finns många celltyper som bär mikrovilli, men de finns oftare i epitelceller, som tarm enterocyter, invecklad kanal epitel av njurar och epididymus. Mikrovilli kan också hittas i vissa specialiserade sensoriska celler som luktreceptorneuroner, organ av Cortis receptorceller, placenta celler, och i celler som rör sig., Även om mikrovilli visar liknande extern morfologi i olika celltyper, innehåller de distinkt molekylär komposition och visar vissa skillnader i deras interna struktur. Nedan behandlar vi funktionerna i mikrovilli av enterocyter i tunntarmen epitel.
bildning
mikrovilli bildas efter att aktinfilament blir associerade med en tät platta som finns i plasmamembranets cytosoliska yta. Proteiner för att samla aktinfilament i buntar och stabilisera dem behövs också. Det ser ut som om det är en sekventiell process., För det första underlättar villin bildandet av buntar av aktinfilament och stabiliserar deras distala ändar som hämmar glödtråden växer, så villin kan vara viktigt att fastställa längden på mikrovilli. Ezrin, mindre riklig, hjälper till att ansluta aktinfilamenten i buntet till plasmamembranet. En andra fas av mikrovillibildning innefattar att nå den slutliga längden och fördelningen på cellytan som hexagoner. Fimbrin och spin kan bidra till dessa funktioner. Sidoanslutningar av protocadheriner mellan aktinfilament av olika mikrovilli sker också under detta andra steg., Det är viktigt att märka att under mikrovillibildning behövs en stor mängd membran-och membranproteiner, vilket levereras av en intensiv exocytos. Evaginationer av plasmamembran, en per varje mikrovillositet, är en följd av aktinfilamentpolymerisering.
struktur
mikrovilli består av 6 huvudproteiner: aktin, fimbrin, villin, myosin (Myo1A), calmodulin och spektrin (icke erytrocytisk) (Figur 3). Mikrovellositetsskelettet är ett bunt av 30-40 aktinfilament parallellt med den långa axeln och med deras minus ändar mot spetsen av mikrovellositeten., Fimbrin och villin kopplar samman aktinfilamenten, medan Myo1A och calmodulin gör anslutningarna mellan bunten och membranet. Plusänden av aktinfilament sträcker sig in i cytoplasman och där aktinfilamenten av olika mikrovilli intermingle mellan varandra och bildar en bana med ett sexkantigt mönster. Denna webb är känd som terminal web, och det sprider sig genom cell periferin under mikrovilli. Terminal web innehåller stor mängd icke-erytrocytisk spektrin.
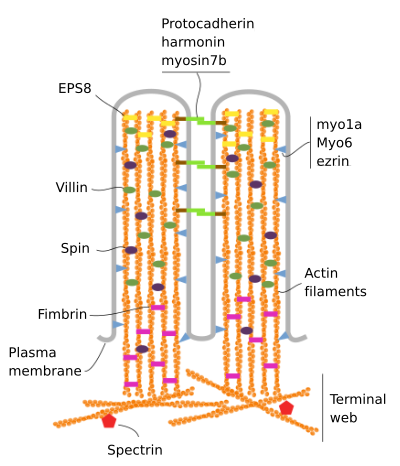
Figur 3., Molekylär organisation av mikrovilli (anpassad från Crawley et al., 2014)
varje mikrovellositet är stabil och rör sig inte, men dess proteinskelett är i permanent förnyelse genom att utbyta aktin och andra byggnadsställningar proteiner mellan mikrovellositeten och cytosolen. Det betyder att det finns en balans mellan inkommande och utgående proteiner. Det uppskattas att varje 20 minuter är skelettet av varje mikrovellositet slutfört förnyat. Mikrovilli kan vara helt oorganiserad och försvinna efter en ökning av kalciumkoncentrationen, till exempel under cellspänning., Kalcium förändrar beteendet hos villin, från stabilisering till avskiljning av aktinfilament. Mikrovilli försvinner i celler som kommer in i mitos. Terminalbanan är också en plaststruktur.
funktion
utbyte av ämnen mellan håligheter eller yttre miljö och vävnader är en av de viktigaste funktionerna i epitel, såsom tarm och njure epitel. Det görs mestadels på epitelcellernas apikala (fria) yta, där de flesta molekylära transportörer, jonpumpar finns och endocytiska processer sker., Ju mer cellyta desto mer utrymme för att rymma molekylära maskiner för utbyte. Mikrovilli är ryggradsliknande utsprång som ökar enormt plasmamembranytan och därmed membranproteininnehållet. Detta är särskilt viktigt i de absorptiva och sekretoriska cellerna i epitel. Mikrovilli kan öka cellytan upp till 100 gånger jämfört med ett platt membran. Enterocytmikrovilli bär många enzymer, som inte är involverade i molekylär utbyte, men i matsmältningen.
mikrovilli kan påverka transduktionsmolekylära vägar., Deras membran fungerar som relativt isolerade domäner med en distinkt uppsättning molekyler, som glukostransportörer, jonpumpar och receptorer. Längden av mikrovilli är väl lämpad för att utföra lokal transduktion av de molekylära vägarna oberoende av cytosolen. Dessutom bildar aktinfilamenten och strukturella proteiner byggnadsställning ett tätt nätverk av molekyler som på något sätt filtrerar eller kontrollerar molekylerna som kommer eller slutar insidan av varje mikrovellositet. Denna byggnadsställning kan också fungera som en temporal lagring för kalcium.,
mikrovillis höga densitet gör en fysisk barriär som skyddar mot patogener. Detta är viktigt för tarmepitel. Dessutom är den stora mängden membran som lagras i mikrovilli en reservoar som kan motstå hypertoniska förolämpningar och förhindra att plasmamembranet bryts.
Stereocilia är specialiserade mikrovillositeter som utför sensoriska funktioner. Trots namnet stereocilium, de är faktiskt modifierade microvillosities omvandlas till sensoriska strukturer, och vissa författare föreslår att namnet estereomicrovellosity istället., De finns i epididymisepitelet och i innerörat, som arbetar som mekanoreceptorer som känner av den omgivande vätskerörelsen. Sterocilia av däggdjurets inre öra finns som en del av Cortis organ. De är 10 till 50 µm långa och innehåller mer än 3000 aktinfilament inuti. Ljudvågorna i luften omvandlas i vätskevågor i örat, som stereocilia känner och omvandlas till elektriska signaler som färdas via hörselnerven till hjärnan. Stereocilia är som antenner som tar emot externa signaler, som överförs i elektrisk information.,
vissa mikrovellositeter är specialiserade på avkänning av ljus. Fotoreceptorer är lätta sensoriska celler som finns i visuella organ som näthinnan. De cellulära sensoriska strukturerna är faktiskt modifierade cilia eller mikrovellositeter. Fotoreceptorer med modifierad mikrovellositet för ljusdetektering kan hittas i ryggradslösa visuella strukturer. Dessa fotoreceptorer innehåller mikrovellositeter ordnade i de så kallade rabdomererna. Varje mikrovellositet innehåller visuella pigment som kan reagera på låga ljusintensiteter och är effektiva vid höga ljusintensiteter., Mikrovellositeter organiserar bildandet av den så kallade rhabdomere, och transduktionen molekylär väg gör mikrovillositetsbaserad ljussvar mer förnuftig än cilium ljus sensoriska system av ryggradsdjur.
ett stort antal extracellulära vesiklar föreslås frigöras från mikrovilli. Spetsarna på enterocytmikrovilli kan frigöra vesiklar till omgivningen., Sambandet mellan plasmamembranet och aktin-myosin-nätverket kan bidra till att producera dessa vesiklar genom att dra membran mot den distala delen av varje mikrovellositet, där små delar av membran omvandlas i vesiklar. Dessa extracellulära vesiklar innehåller enzymer och berikas i alkaliskt fosfatas.
bibliografi
brun J W, McKnight C J. 2010. Molekylär modell av microvillar cytoskeleton och organisation av borst gränsen. PLoS One. 5: e940 ![]() .
.
Crawley SW, Mooseker MS, Tyska MJ. 2014. Formar tarmborstgränsen., J Cell Biol. 207: 441-451 ![]() .
.
Fain G L, Hardie R, S Laughlin B. 2010. Fototransduktion och utvecklingen av fotoreceptorer. Curr Biol. 20: R114-R124 ![]() .
.
Lange K. 2011. Mikrovillis grundläggande roll i huvudfunktionerna hos differentierade celler: kontur av ett universellt reglerande och signaleringssystem vid cellens periferi. J Cell Physiol. 226: 896-92
McConnell R E, Higginbotham J N, Shifrin Jr D A, Tabb D L, Coffey R J, Tyska M J. 2009. Enterocyte microvillus är en vesikelgenererande organell. J Cell Biol., 185: 1285-129 ![]() .
.
Sauvanet C, Wayt J, Pelaseyed T, Bretscher A. 2015. Struktur, reglering och funktionell mångfald av mikrovilli på den apikala domänen av epitelceller. Årlig översyn av cell-och utvecklingsbiologi. 31: 593-621.