1. Formacja
2. Struktura
3. Function
Microvilli to krótkie, cienkie wypukłości zwykle występujące na wolnych powierzchniach niektórych zróżnicowanych komórek, częściej w komórkach nabłonkowych (ryc. 1 i 2). Są to struktury nitkowate o długości około 1-2 µm i grubości około 100 nm. Wewnątrz każda mikrowłóknina zawiera dziesiątki włókien aktynowych zorientowanych równolegle do długiej osi., Microvilli są zwykle gęste i szczelnie upakowane jak brush border. W widoku z góry microvilli są przestrzennie zorganizowane jako sześciokąty.

Rysunek 1. Nabłonek jelita cienkiego w mikroskopii świetlnej (obraz po lewej stronie) i skaningowej mikroskopii elektronowej (obraz po prawej stronie) pokazujący mikrowilli na wolnej powierzchni komórek.
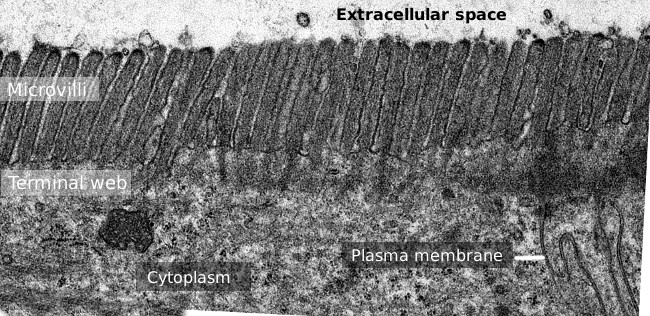
Rysunek 2. Transmisja obraz mikroskopii elektronowej powierzchni nabłonka jelita cienkiego., Taśma końcowa włókien aktynowych jest ciemniejszym pasem obserwowanym od podstaw do microvilli.
istnieje wiele rodzajów komórek noszących microvilli, ale częściej występują one w komórkach nabłonkowych, takich jak enterocyty jelitowe, nabłonek kanałowy nerek i najądrza. Microvilli można również znaleźć w niektórych wyspecjalizowanych komórkach czuciowych, takich jak neurony receptora węchowego, narząd komórek receptora Cortiego, komórki łożyska i w komórkach, które się poruszają., Chociaż microvilli wykazują podobną morfologię zewnętrzną w różnych typach komórek, zawierają odrębny skład molekularny i wykazują pewne różnice w ich strukturze wewnętrznej. Poniżej mamy do czynienia z cechami microvilli enterocytów nabłonka jelita cienkiego.
powstawanie
mikrowłókna powstają po związaniu się włókien aktynowych z gęstą płytką znajdującą się na powierzchni cytozolowej błony plazmatycznej. Potrzebne są również białka do gromadzenia włókien aktynowych w wiązkach i ich stabilizacji. Wygląda na to, że jest to proces sekwencyjny., Po pierwsze, villin ułatwia tworzenie wiązek włókien aktynowych i stabilizuje ich dystalne końce hamując wzrost włókien, więc villin może być ważny dla ustalenia długości microvilli. Ezrin, mniej obfity, pomaga połączyć włókna aktynowe wiązki z błoną osocza. Druga faza powstawania mikrowilli obejmuje osiągnięcie końcowej długości i rozmieszczenie na powierzchni komórki w postaci sześciokątów. Fimbrin i spin mogą przyczynić się do tych cech. Boczne połączenia protocadherins między aktyny włókien różnych microvilli również zdarzają się w tym drugim etapie., Ważne jest, aby zauważyć, że podczas tworzenia microvilli potrzebna jest duża ilość błon i białek błonowych, które są dostarczane przez intensywną egzocytozę. Ewaginacja błony plazmowej, po jednej na każdą mikrowilość, jest konsekwencją polimeryzacji włókien aktynowych.
struktura
Microvilli składa się z 6 głównych białek: aktyny, fimbryny, villina, miozyny (Myo1A), kalmoduliny i spektryny (nie erytrocyty) (ryc. 3). Szkielet microvellosity jest wiązką 30-40 włókien aktynowych równoległych do długiej osi i z ich minus końcami w kierunku wierzchołka mikrowellosity., Fimbrin i villin łączą ze sobą włókna aktyny, podczas gdy Myo1A i kalmodulina tworzą połączenia między wiązką a błoną. Plus koniec włókien aktynowych rozciąga się do cytoplazmy i tam włókna aktynowe różnych mikrowłókienek mieszają się między sobą i tworzą sieć o sześciokątnym wzorze. Sieć ta jest znana jako sieć terminalna i rozprzestrzenia się przez obrzeża komórkowe Pod microvilli. Wstęga końcowa zawiera dużą ilość spektryny nie erytrocytowej.
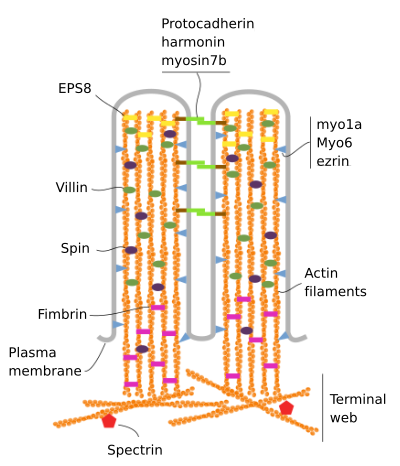
Rysunek 3., Molekularna organizacja microvilli (zaadaptowana z Crawley et al., 2014)
każda mikrowelastyczność jest stabilna i nie porusza się, ale jej szkielet białkowy jest trwale odnawiany przez wymianę aktyny i innych białek rusztowania między mikrowelastycznością a cytozolem. Oznacza to, że istnieje równowaga między przychodzącymi i wychodzącymi białkami. Szacuje się, że co 20 minut szkielet każdej mikrowłókniny jest odnawiany. Microvilli może być całkowicie zdezorganizowany i zanikać po zwiększeniu stężenia wapnia, na przykład pod wpływem stresu komórkowego., Wapń zmienia zachowanie villina, od stabilizującego do odcinającego włókna aktyny. Microvilli znikają w komórkach wchodzących w mitozę. Taśma zaciskowa jest również konstrukcją z tworzywa sztucznego.
funkcja
wymiana substancji między jamami lub środowiskiem zewnętrznym a tkankami jest jedną z głównych funkcji nabłonka, takich jak nabłonek jelita i nerki. Odbywa się głównie na wierzchołkowej (wolnej) powierzchni komórek nabłonkowych, gdzie znajduje się większość transporterów molekularnych, pomp jonowych i zachodzą procesy endocytarne., Im więcej powierzchni komórki, tym więcej miejsca na urządzenie molekularne do wymiany. Microvilli to występy przypominające kręgosłup, które znacznie zwiększają powierzchnię błony plazmowej, a tym samym zawartość białka w błonie. To szczególnie ważne w tych chłonnych i wydzielniczych komórek nabłonka. Microvilli może zwiększyć powierzchnię komórki do 100 razy w porównaniu z płaską błoną. Mikrocyty enterocytowe noszą wiele enzymów, które nie biorą udziału w wymianie molekularnej, ale w trawieniu.
Microvilli może wpływać na transdukcyjne szlaki molekularne., Ich błony działają jako względnie izolowane domeny z odrębnym zestawem cząsteczek, takich jak transportery glukozy, pompy jonowe i receptory. Długość microvilli jest dobrze przystosowana do wykonywania lokalnych transdukcji szlaków molekularnych niezależnie od cytozolu. Ponadto włókna aktynowe i strukturalne białka tworzą gęstą sieć cząsteczek, które w jakiś sposób filtrują lub kontrolują cząsteczki wchodzące lub wychodzące z wnętrza każdej mikrowłókniny. Rusztowanie to może również służyć jako czasowe magazynowanie wapnia.,
wysoka gęstość microvilli tworzy barierę fizyczną chroniącą przed patogenami. Jest to ważne dla nabłonka jelita. Ponadto duża ilość membrany przechowywana w microvilli jest zbiornikiem, który może wytrzymać hipertoniczne obelgi i zapobiegać pękaniu błony plazmowej.
Stereocilia są wyspecjalizowanymi mikrocząsteczkami pełniącymi funkcje sensoryczne. Pomimo nazwy stereocilium, są to w rzeczywistości zmodyfikowane mikrowilosity przekształcone w struktury czuciowe, a niektórzy autorzy proponują zamiast nich nazwę estereomicrovellosity., Można je znaleźć w nabłonku najądrza oraz w uchu wewnętrznym, pracując jako mechanoreceptory, które wyczuwają otaczający ruch płynu. Sterocilia ucha wewnętrznego ssaka występuje jako część narządu Cortiego. Mają od 10 do 50 µm długości i zawierają ponad 3000 włókien aktynowych w środku. Fale dźwiękowe powietrza są przekształcane w płynne fale w uchu, które stereocylia wyczuwają i przekształcają się w sygnały elektryczne, które podróżują przez nerw słuchowy do mózgu. Stereocilia są jak anteny, które odbierają sygnały zewnętrzne, które są przekazywane w informacji elektrycznej.,
niektóre mikroelementy specjalizują się w wykrywaniu światła. Fotorreceptory to lekkie komórki czuciowe znajdujące się w narządach wzrokowych, takich jak siatkówka. Komórkowe struktury czuciowe są w rzeczywistości zmodyfikowanymi rzęskami lub mikrowłóknami. Fotoreceptory o zmodyfikowanych mikroelementach do wykrywania światła można znaleźć w strukturach wizualnych bezkręgowców. Fotoreceptory te zawierają mikroelementy ułożone w tzw. rabdomerach. Każda mikrocząsteczka zawiera pigmenty wizualne, które mogą reagować na słabe natężenie światła i są wydajne przy wysokiej intensywności światła., Mikrowłókniny organizują się tworząc tzw. rabdomerę, a szlak molekularny transdukcji sprawia, że mikrowłóknikowatość oparta na światłach jest bardziej sensowna niż światłowodowe systemy sensoryczne kręgowców.
proponuje się uwolnienie dużej liczby pęcherzyków zewnątrzkomórkowych z microvilli. Końcówki enterocyte microvilli są w stanie uwolnić pęcherzyki do otaczającego środowiska., Połączenie między błoną plazmową a siecią aktyny-miozyny może pomóc w wytworzeniu tych pęcherzyków, przeciągając błonę w kierunku dystalnej części każdej mikrowłókniny, gdzie małe części błony przekształcają się w pęcherzyki. Te pozakomórkowe pęcherzyki zawierają enzymy i są wzbogacone w fosfatazę alkaliczną.
Bibliografia
Brown J W, McKnight C J. 2010. Model molekularny cytoszkieletu mikrowilarnego i organizacji granicy szczotki. PLoS 1. 5: e940 ![]() .
.
Crawley SW, Mooseker MS, Tyska MJ. 2014. Kształtowanie granicy pędzla jelitowego., J Cell Biol. 207: 441-451 ![]() .
.
Fain G L, Hardie R, Laughlin s B. 2010. Fototransdukcji i ewolucji fotoreceptorów. Curr Biol. 20: R114-R124 ![]() .
.
Lange K. 2011. Podstawowa rola microvilli w głównych funkcjach zróżnicowanych komórek: zarys uniwersalnego systemu regulacji i sygnalizacji na obwodzie komórki. J Cell Physiol. 226: 896-92
McConnell R E, Higginbotham J N, Shifrin Jr D A, Tabb D L, Coffey R J, Tyska M J. 2009. Enterocyte microvillus to organelle wytwarzające pęcherzyki. J Cell Biol., 185: 1285-129 ![]() .
.
Sauvanet C, Wayt J, pelas T, Bretscher A. 2015. Struktura, Regulacja i funkcjonalna różnorodność microvilli na wierzchołkowej domenie nabłonka komórki. Annual review of cell and development biology. 31: 593-621.