1. Formáció
2. Szerkezet
3. Funkció
Microvilli rövid vékony kiálló általában a szabad felületek néhány differenciált sejtek, gyakoribb a hámsejtek (Számok 1, 2). Ezek körülbelül 1-2 µm hosszú és körülbelül 100 nm vastag filiform szerkezetek. Belül minden mikrovillozitás több tucat aktinszálat tartalmaz, amelyek a hosszú tengelygel párhuzamosan vannak orientálva., A mikrovillák általában sűrűek, szorosan csomagolva, mint egy kefe szegély. Felülnézetben a mikrovillák térben hatszögekként vannak elrendezve.

1.ábra. Vékonybél epithelium fénymikroszkóppal (kép a bal oldalon) és pásztázó elektronmikroszkóppal (kép a jobb oldalon), amely mikrovillákat mutat a sejtek szabad felületén.
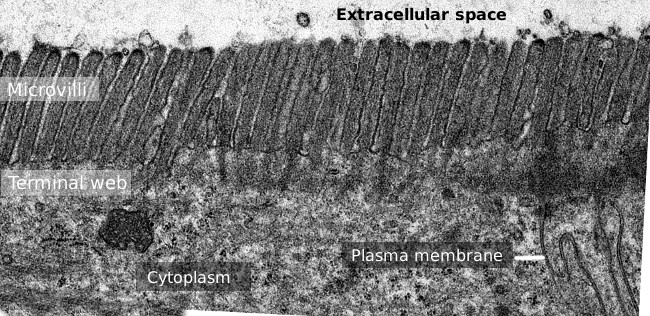
2.ábra. A vékonybél epithelium felületének átviteli elektronmikroszkópos képe., Az aktin filamentumok terminális hálója a sötétebb sáv, amelyet a mikrovillák észlelnek.
sok cella típusok csapágy microvilli, de gyakrabban megtalálható a hámsejtek, mint gut enterocytes, szövevényes csatorna hám vese, epididymus. A mikrovillák néhány speciális szenzoros sejtben is megtalálhatók, mint például a szaglóreceptor neuronok, a Corti receptor sejtjeinek szervei, a placenta sejtek, valamint a mozgó sejtekben., Bár a mikrovillák hasonló külső morfológiát mutatnak különböző sejttípusokban, különálló molekuláris összetételt tartalmaznak, és belső szerkezetükben némi eltérést mutatnak. Az alábbiakban a vékonybél epitéliumának enterocitáinak mikrovilláinak jellemzőivel foglalkozunk.
formáció
a mikrovillák azután alakulnak ki, hogy az aktinszálak a plazmamembrán citoszolos felületén található sűrű lemezhez kapcsolódnak. Az aktinszálak kötegekben történő összegyűjtésére és stabilizálására szolgáló fehérjékre is szükség van. Úgy tűnik, ez egy szekvenciális folyamat., Először is, a villin elősegíti az aktinszálak kötegeinek kialakulását, és stabilizálja az izzószál növekedését gátló disztális végeiket, így a villin fontos lehet a mikrovillák hosszának megállapításához. A kevésbé bőséges Ezrin segít a köteg aktinszálainak a plazmamembránhoz való csatlakoztatásában. A mikrovillák képződésének második fázisa magában foglalja a végső hossz elérését, valamint a sejtfelszín hatszögként való eloszlását. A Fimbrin és a spin hozzájárulhat ezekhez a funkciókhoz. A protocadherinek oldalirányú összeköttetései a különböző mikrovillák aktinszálai között szintén előfordulnak ebben a második szakaszban., Fontos megjegyezni, hogy a mikrovillák képződése során nagy mennyiségű membrán-és membránfehérjére van szükség, amelyet intenzív exocitózis biztosít. A plazmamembrán evaginációi, minden egyes mikrovillozitásonként egy, az aktin szálas polimerizáció következménye.
szerkezet
a mikrovillák 6 fő fehérjéből állnak: aktin, fimbrin, villin, myosin (Myo1A), calmodulin és spectrin (nem eritrocitikus) (3.ábra). A mikrovellozitású csontváz egy 30-40 aktinszálból álló köteg, amely párhuzamos a hosszú tengellyel, mínusz végükkel a mikrovellozitás csúcsa felé., A Fimbrin és a villin összekapcsolják az aktinszálakat, míg a Myo1A és a calmodulin a köteg és a membrán közötti kapcsolatot hozza létre. Az aktinszálak plusz vége a citoplazmába nyúlik, ott a különböző mikrovillák aktinszálai összekeverednek egymással, és hatszögletű mintázatot alkotnak. Ez a web terminal web néven ismert, és a mikrovillák alatt elterjed a sejt perifériáján. A terminális web nagy mennyiségű nem eritrocitikus spektrint tartalmaz.
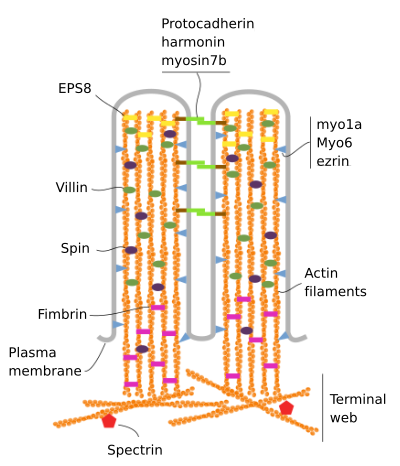
3.ábra., Molekuláris szervezése microvilli (adaptált Crawley et al., 2014)
Minden microvellosity stabil, nem mozog, de a fehérje csontváz állandó megújítása által cseréje aktin, illetve egyéb állvány fehérjék között a microvellosity a cytosol. Ez azt jelenti, hogy egyensúly van a bejövő és a kimenő fehérjék között. Becslések szerint 20 percenként megújul az egyes mikrovellózisok csontváza. A mikrovillák teljesen szervezetlenek lehetnek, és a kalciumkoncentráció növekedése után eltűnhetnek, például sejtes stressz esetén., A kalcium megváltoztatja a villin viselkedését, a stabilizációtól az aktinszálak megszakításáig. A mikrovillák eltűnnek a mitózisba belépő sejtekben. A terminálháló műanyag szerkezet is.
Function
anyagok cseréje üregek vagy külső környezet és szövetek között az epithelia egyik fő funkciója, mint például a bél és a vese epithelia. Többnyire az epiteliális sejtek apikális (szabad) felületén történik, ahol a legtöbb molekuláris transzporter, ionszivattyú található, és endocitikus folyamatok zajlanak., Minél több sejtfelület van, annál több hely van a csere molekuláris gépeinek befogadására. A mikrovillák gerincszerű kiemelkedések, amelyek óriási mértékben növelik a plazmamembrán felületét, ezért a membrán fehérjetartalmát. Ez különösen fontos az epithelia abszorpciós és szekréciós sejtjeiben. A mikrovillák akár 100-szor is növelhetik a sejtfelületet a lapos membránhoz képest. Az enterocita mikrovillák számos enzimet hordoznak, amelyek nem vesznek részt a molekuláris cserében, hanem az emésztésben.
a mikrovillák befolyásolhatják a transzdukciós molekuláris útvonalakat., Membránjaik viszonylag elszigetelt doménekként működnek, különböző molekulákkal, például glükózszállítókkal, ionszivattyúkkal és receptorokkal. A mikrovillák hossza kiválóan alkalmas a molekuláris útvonalak lokális transzdukciójának elvégzésére a citozoltól függetlenül. Ezenkívül az aktinszálak és a szerkezeti fehérjék sűrű molekulahálózatot alkotnak, amely valahogy kiszűri vagy szabályozza az egyes mikrovellózisok belsejéből érkező vagy kilépő molekulákat. Ez az állvány a kalcium időbeli tárolásaként is működhet.,
a mikrovillák nagy sűrűsége fizikai akadályt képez, amely védelmet nyújt a kórokozókkal szemben. Ez fontos a bélhám esetében. Ezenkívül a mikrovillákban tárolt nagy mennyiségű membrán olyan tartály, amely ellenáll a hipertóniás sértéseknek, megakadályozza a plazmamembrán törését.
A Stereocilia szenzoros funkciókat ellátó speciális mikrovillositások. A stereocilium név ellenére valójában átalakított mikrovillositások, amelyek szenzoros struktúrákká alakulnak át, egyes szerzők inkább az estereomicrovellosity nevet javasolják., Ezek megtalálhatók az epididymis epitéliumban, a belső fülben pedig mechanoreceptorokként működnek, amelyek érzékelik a környező folyadékmozgást. Az emlős belső fülének sterociliája a Corti szervének részeként található. 10-50 µm hosszúak, és több mint 3000 aktinszálat tartalmaznak. A hanghullámok a levegő át a folyadék a hullámok a fül, amely sztereociliumok értelemben pedig át elektromos jelekké utazik keresztül hallóideg, hogy az agy. A sztereocília olyan antennák, amelyek külső jeleket kapnak, amelyek elektromos információkban vannak átalakítva.,
néhány mikrovellózis a fény érzékelésére szakosodott. A fotoreceptorok olyan könnyű érzékszervi sejtek, amelyek a vizuális szervekben, például a retinában találhatók. A celluláris szenzoros struktúrák valójában módosított csillók vagy mikrovellózisok. A fényérzékeléshez módosított mikrovellózisokat hordozó fotoreceptorok gerinctelen vizuális struktúrákban találhatók. Ezek a fotoreceptorok az úgynevezett rhabdomerekben elrendezett mikrovellozitásokat tartalmazzák. Minden mikrovellozitás olyan vizuális pigmenteket tartalmaz, amelyek képesek reagálni az alacsony fényerősségre, és nagy fényerősség esetén hatékonyak., A mikrovellózisok az úgynevezett rhabdomere kialakulását szervezik, a transzdukciós molekuláris út pedig a mikrovillozitás alapú fényválaszt érzékenyebbé teszi, mint a gerincesek cilium fényérzékelő rendszerei.
számos extracelluláris vezikulumot javasolnak felszabadítani a mikrovillákból. Az enterocita mikrovillák tippjei képesek a hólyagok felszabadítására a környező környezetbe., A plazmamembrán és az aktin-myosin hálózat közötti kapcsolat segíthet ezen hólyagok előállításában azáltal, hogy a membránt az egyes mikrovellozitások disztális része felé húzza, ahol a membrán kis részei hólyagokban alakulnak át. Ezek az extracelluláris vezikulumok enzimeket tartalmaznak, alkalikus foszfatázban dúsítva.
bibliográfia
Brown J W, McKnight C J. 2010. A mikrovilláris citoszkeleton molekuláris modellje, valamint az ecset határának megszervezése. PLoS One. 5: e940 ![]() .
.
Crawley SW, Mooseker MS, Tyska MJ. 2014. A bélkefe határának kialakítása., J Cell Biol. 207: 441-451 ![]() .
.
Fain G L, Hardie R, Laughlin s B. 2010. A fototranszdukció és a fotoreceptorok fejlődése. Curr Biol. 20: R114-R124 ![]() .
.
Lange K. 2011. A mikrovillák alapvető szerepe a differenciált sejtek fő funkcióiban: egy univerzális szabályozó-jelző rendszer vázlata a sejt perifériáján. J Sejt Physiol. 226: 896-92
McConnell R E, Higginbotham J N, Shifrin Jr D A, Tabb D L, Coffey R J, Tyska M J. 2009. Az enterocita microvillus egy vezikulumtermelő organelle. J Cell Biol., 185: 1285-129 ![]() .
.
Sauvanet C, Wayt J, Pelasyed T, Bretscher A. 2015. A mikrovillák szerkezete, szabályozása és funkcionális sokfélesége az epitheliális sejtek apikális tartományában. A sejt-és fejlődésbiológia éves áttekintése. 31: 593-621.