1. Formarea
2. Structura
3. Funcția
Microvilli sunt proeminențe scurte subțiri care se găsesc de obicei în suprafețele libere ale unor celule diferențiate, mai frecvente în celulele epiteliale (Figurile 1 și 2). Sunt structuri filiforme de aproximativ 1 până la 2 µm lungime și aproximativ 100 nm grosime. În interior, fiecare microvilozitate conține zeci de filamente de actină orientate paralel cu axa lungă., Microvilli sunt de obicei densi și bine împachetați ca o margine de perie. În vedere de sus, microvilli sunt organizate spațial ca hexagoane.

Figura 1. Epiteliu intestinal mic la microscopia luminoasă (imaginea din stânga) și la microscopia electronică de scanare (imaginea din dreapta) care prezintă microvilli la suprafața liberă a celulelor.
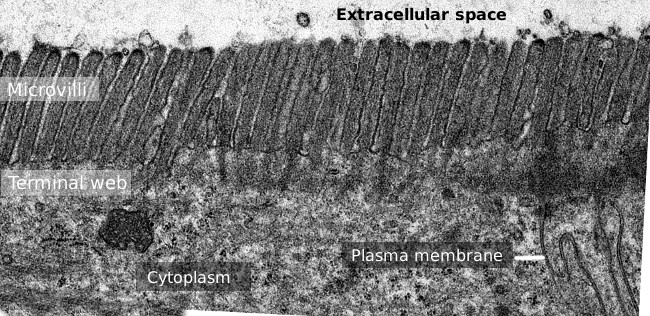
Figura 2. Imagine microscopie electronică de transmisie a suprafeței epiteliului intestinului subțire., Pânza terminală a filamentelor de actină este banda mai întunecată observată bazal la microvilli.există multe tipuri de celule care poartă microvilli, dar se găsesc mai frecvent în celulele epiteliale, cum ar fi enterocitele intestinale, epiteliul duct convolut al rinichilor și epididimul. Microvilli pot fi, de asemenea, găsite în unele celule senzoriale specializate, cum ar fi neuronii receptorilor olfactivi, organul celulelor receptorilor Corti, celulele placentei și în celulele care se mișcă., Deși microvilli prezintă morfologie externă similară în diferite tipuri de celule, ele conțin o compoziție moleculară distinctă și prezintă unele diferențe în structura lor internă. Mai jos, avem de-a face cu caracteristicile microvililor enterocitelor epiteliului intestinului subțire.Microvilli se formează după ce filamentele de actină se asociază cu o placă densă găsită pe suprafața citosolică a membranei plasmatice. Proteinele pentru colectarea filamentelor de actină în mănunchiuri și stabilizarea acestora sunt, de asemenea, necesare. Se pare că este un proces secvențial., În primul rând, villin facilitează formarea de mănunchiuri de filamente de actina si stabilizeaza lor distală se termină inhibarea filament crească, astfel încât villin poate fi important să se stabilească lungimea de microvili. Ezrin, mai puțin abundent, ajută la conectarea filamentelor de actină ale mănunchiului la membrana plasmatică. O a doua fază de formare a microvililor include atingerea lungimii finale și distribuția pe suprafața celulei sub formă de hexagoane. Fimbrin și spin pot contribui la aceste caracteristici. Conexiunile laterale prin protocadherine între filamentele de actină ale diferitelor microvilli se întâmplă și în această a doua etapă., Este important de observat că în timpul formării microvililor este necesară o cantitate mare de proteine membranare și membrane, care este furnizată de o exocitoză intensă. Evaginările membranei plasmatice, câte una pentru fiecare microvilozitate, sunt o consecință a polimerizării filamentului de actină.
Structura
Microvililor sunt alcătuite din 6 proteine principale: actina, fimbrin, villin, miozina (Myo1A), calmodulin și spectrină (non eritrocitar) (Figura 3). Scheletul de microvelozitate este un pachet de 30-40 filamente de actină paralele cu axa lungă și cu capetele lor minus spre vârful microvelozității., Fimbrin și villin leagă împreună filamentele de actină, întrucât Myo1A și calmodulin face conexiuni între pachet și membrană. Capătul plus al filamentelor de actină se extinde în citoplasmă și acolo filamentele de actină ale diferitelor microvilli se amestecă între ele și formează o rețea cu un model hexagonal. Acest web este cunoscut sub numele de terminal web, și se răspândește prin periferia celulară sub microvilli. Terminalul web conține o cantitate mare de spectrină non-eritrocitară.
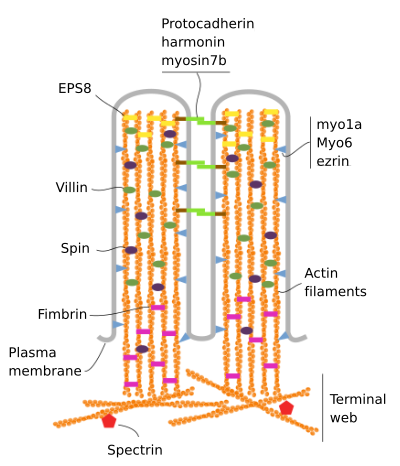
Figura 3., Organizarea moleculară a microvilli (adaptat de la Crawley și colab., 2014)
Fiecare microvellosity este stabilă și nu se mișcă, dar proteine de schelet este în permanentă reînnoire prin schimbul de actina și alte schele proteine între microvellosity și citosol. Aceasta înseamnă că există un echilibru între proteinele de intrare și cele de ieșire. Se estimează că la fiecare 20 de minute scheletul fiecărei microvelozități este completat reînnoit. Microvilli pot fi complet dezorganizați și dispar după o creștere a concentrației de calciu, de exemplu sub stres celular., Calciul schimbă comportamentul vilinei, de la stabilizarea la ruperea filamentelor de actină. Microvilli dispar în celulele care intră în mitoză. Pânza terminalului este, de asemenea, o structură din plastic.
Funcția
Schimbul de substanțe între cavități sau mediul extern și țesuturi este una funcția principală a epiteliilor, cum ar fi intestin și rinichi epitelii. Se face în cea mai mare parte la suprafața apicală (liberă) a celulelor epiteliale, unde se află majoritatea transportorului molecular, pompele de ioni și se întâmplă procese endocitare., Cu cât suprafața celulară este mai mare, cu atât mai mult spațiu pentru a găzdui mașinile moleculare pentru schimb. Microvilli sunt proeminențe asemănătoare coloanei vertebrale care cresc enorm suprafața membranei plasmatice și, prin urmare, conținutul de proteine din membrană. Acest lucru este deosebit de important în acele celule absorbante și secretoare ale epiteliilor. Microvilli poate crește suprafața celulei de până la 100 de ori în comparație cu o membrană plană. Microvilli enterocite poartă multe enzime, care nu sunt implicate în schimbul molecular, ci în digestie.Microvilli pot influența căile moleculare de transducție., Membranele lor funcționează ca domenii relativ izolate, cu un set distinct de molecule, cum ar fi transportorii de glucoză, pompele de ioni și receptorii. Lungimea microvililor este potrivită pentru efectuarea transducției locale a căilor moleculare independent de citozol. În plus, filamentele de actină și proteinele structurale formează o rețea densă de molecule care filtrează sau controlează cumva moleculele care vin sau renunță la interiorul fiecărei microvelozități. Această schelă poate funcționa și ca un depozit temporal pentru calciu.,densitatea mare a microvillilor face o barieră fizică care protejează împotriva agenților patogeni. Acest lucru este important pentru epiteliul intestinal. În plus, cantitatea mare de membrană stocată în microvilli este un rezervor care poate rezista insultelor hipertonice și poate preveni ruperea membranei plasmatice. Stereocilia sunt microvilozități specializate care îndeplinesc funcții senzoriale. În ciuda numelui stereocilium, ele sunt de fapt modificate microvillosities transformate în structuri senzoriale, și unii autori propun numele estereomicrovellosity în loc., Ele pot fi găsite în epiteliul epididimului și în urechea internă, funcționând ca mecanoreceptori care simt mișcarea fluidului din jur. Sterocilia urechii interne a mamiferelor se găsește ca parte a organului Corti. Acestea au o lungime de 10 până la 50 µm și conțin mai mult de 3000 de filamente de actină în interior. Undele sonore ale aerului sunt transformate în unde fluide în ureche, pe care stereocilia le simt și se transformă în semnale electrice care circulă prin nervul auditiv către creier. Stereocilia sunt ca antene care primesc semnale externe, care sunt transduse în informații electrice., unele microvelozități sunt specializate în detectarea luminii. Fotorreceptorii sunt celule senzoriale ușoare care se găsesc în organele vizuale precum retina. Structurile senzoriale celulare sunt de fapt modificate cilia sau microvelozități. Fotoreceptorii care poartă microvelozități modificate pentru detectarea luminii pot fi găsite în structurile vizuale nevertebrate. Acești fotoreceptori conțin microvelozități aranjate în așa-numitele rabdomere. Fiecare microvelozitate conține pigmenți vizuali care pot răspunde la intensități reduse ale luminii și sunt eficienți la intensități ridicate ale luminii., Microvellosities organiza formând așa-numitele rhabdomere, și transducție cale moleculară face microvillosity bazate pe lumină răspuns mai sensibil decât cilium lumina sisteme senzoriale de vertebrate.un număr mare de vezicule extracelulare sunt propuse pentru a fi eliberate din microvilli. Sfaturile microvililor enterocite sunt capabile să elibereze vezicule în mediul înconjurător., Legătura dintre membrana plasmatică și rețeaua actină-miozină poate ajuta la producerea acestor vezicule prin tragerea membranei spre partea distală a fiecărei microvelozități, unde porțiuni mici de membrană sunt transformate în vezicule. Aceste vezicule extracelulare conțin enzime și sunt îmbogățite în fosfatază alcalină.
Bibliografie
Brown J W, McKnight C J. 2010. Modelul Molecular al citoscheletului microvilar și organizarea marginii periei. PLoS Unu. 5: e940 ![]() . Crawley SW, Mooseker MS, Tyska MJ. 2014. Modelarea marginii periei intestinale., Biol Al Celulei J. 207: 441-451
. Crawley SW, Mooseker MS, Tyska MJ. 2014. Modelarea marginii periei intestinale., Biol Al Celulei J. 207: 441-451 ![]() .
.
Fain G L, Hardie R, Laughlin S B. 2010. Fototransducția și evoluția fotoreceptorilor. Curr Biol. 20: R114-R124 ![]() .Lange K. 2011. Rolul Fundamental al microvillilor în principalele funcții ale celulelor diferențiate: conturul unui sistem universal de reglare și semnalizare la periferia celulei. Physiol Al Celulei J. 226: 896-92
.Lange K. 2011. Rolul Fundamental al microvillilor în principalele funcții ale celulelor diferențiate: conturul unui sistem universal de reglare și semnalizare la periferia celulei. Physiol Al Celulei J. 226: 896-92
McConnell R E, Higginbotham J N, Shifrin Jr D A, Tabb D L, Coffey R J, Tyska M J. 2009. Microvilul enterocit este o organelle generatoare de vezicule. Biol Al Celulei J., 185: 1285-129 ![]() .sauvanet C, Wayt J, Pelaseyed T, Bretscher A. 2015. Structura, reglarea și diversitatea funcțională a microvililor pe domeniul apical al celulelor epiteliale. Revizuirea anuală a biologiei celulare și de dezvoltare. 31: 593-621.
.sauvanet C, Wayt J, Pelaseyed T, Bretscher A. 2015. Structura, reglarea și diversitatea funcțională a microvililor pe domeniul apical al celulelor epiteliale. Revizuirea anuală a biologiei celulare și de dezvoltare. 31: 593-621.