– 1. Dannelse
2. Struktur
3. Funksjon
Microvilli er korte tynne utspring vanligvis finnes i gratis overflater av noen differensierte celler, mer hyppige i epitelceller (Figurene 1 og 2). De er filiform strukturer på ca 1 til 2 µm lange og rundt 100 nm tykk. Inne i hver microvillosity inneholder dusinvis av actin filamenter orientert parallelt med den lange aksen., Microvilli er vanligvis tett og tett pakket som en pensel grensen. I topp utsikt, microvilli er romlig organisert som sekskanter.

Figur 1. Små tarmens epitel på lys-mikroskopi (bildet til venstre) og scanning elektron mikroskopi (bildet til høyre) som viser microvilli på den frie overflaten av cellene.
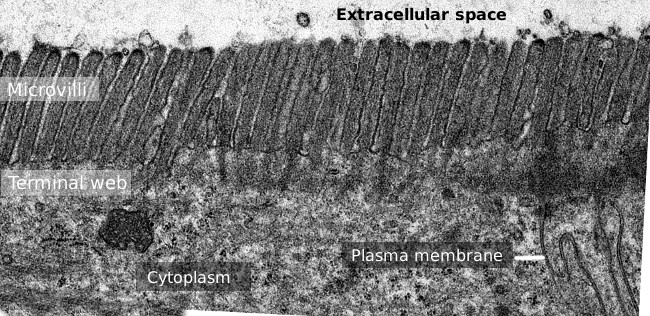
Figur 2. Transmisjon elektron mikroskopi bilde av tynntarmen epitel overflaten., Terminal web av actin filamenter er mørkere band observert basal å microvilli.
Det er mange celletyper lager microvilli, men de er oftere funnet i epitelceller, som gut enterocytes, convoluted duct epitel av nyrer og epididymus. Microvilli kan også bli funnet i noen spesialiserte sensoriske celler som olfactory reseptor nevroner, orgel av Corti ‘ s reseptor celler, morkaken celler, og i celler som er i bevegelse., Selv om microvilli vis lignende ytre morfologi i forskjellige celletyper, de inneholder forskjellige molekylære sammensetning og vise noen forskjeller i deres interne struktur. Nedenfor har vi å gjøre med funksjoner av microvilli av enterocytes av tynntarmen epitel.
Dannelse
Microvilli er dannet etter actin filamenter få forbundet med en tett plate funnet i cytosolic overflaten av plasma membran. Proteiner for å samle actin filamenter i bunter og stabiliserende dem er også nødvendig. Det ser ut som det er en sekvensiell prosess., Første, villin muliggjør dannelsen av bunter av actin filamenter og stabiliserer deres distale ender hemme filament vokse, så villin kan være viktig å etablere lengden på microvilli. Ezrin, mindre rikelig, bidrar til å koble actin filamenter av pakken til plasma membran. En annen fase av microvilli dannelse har nå endelig lengde og fordeling på celleoverflaten som sekskanter. Fimbrin og spinn kan bidra til disse funksjonene. Lateral forbindelser med protocadherins mellom actin filamenter av ulike microvilli også skje i løpet av denne andre fasen., Det er viktig å merke seg at under microvilli dannelsen av en stor mengde membran og membranproteiner er nødvendig, som er levert av en intens exocytosis. Evaginations av plasma membran, en for hver microvillosity, er en konsekvens av actin filament polymerisering.
– Struktur
Microvilli består av 6 viktigste proteiner: actin, fimbrin, villin, myosin (Myo1A), calmodulin og spectrin (ikke erythrocytic) (Figur 3). Den microvellosity skjelettet er en bunt av 30-40 actin filamenter parallell til den lange aksen og med sine minus ender mot spissen av microvellosity., Fimbrin og villin knytte sammen de actin filamenter, mens Myo1A og calmodulin gjøre tilkoblinger mellom pakke og membranen. Pluss-enden av actin filamenter strekker seg inn i cytoplasma og det actin filamenter av ulike microvilli blander mellom hverandre og danne et nett med et sekskantet mønster. Denne web er kjent som terminal web, og det sprer seg gjennom cellen periferi under microvilli. Terminal web inneholder store mengder ikke-erythrocytic spectrin.
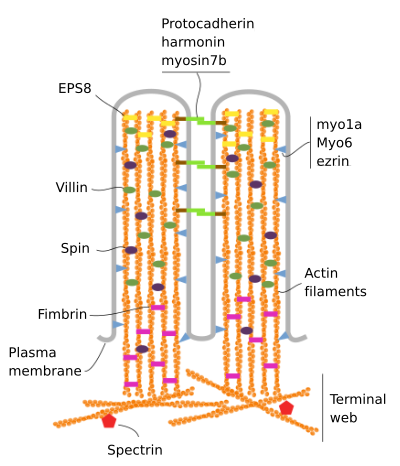
Figur 3., Molekylær organisering av microvilli (tilpasset fra Crawley et al., 2014)
Hver microvellosity er stabil og ikke flytte, men dens protein skjelettet er i kontinuerlig fornyelse av utveksling av actin og andre stillaset proteiner mellom microvellosity og cytosol. Det betyr at det er en balanse mellom inngående og utgående proteiner. Det er anslått at hver 20 minutter skjelettet av hver microvellosity er ferdig fornyet. Microvilli kan være helt uorganisert og forsvinner etter en økning i kalsium-konsentrasjon, for eksempel under celle stress., Kalsium endrer atferden til villin, fra stabiliserende til kutting actin filamenter. Microvilli forsvinne i cellene inn mitose. Terminalen nettet er også en plast struktur.
Funksjon
Utveksling av stoffer mellom hulrom eller ytre miljø og vev er en av de viktigste funksjonen av epithelia, for eksempel tarmen og nyrene epithelia. Det er for det meste gjort på apical (gratis) overflaten av epiteliale celler, der de fleste molekylær transporter, ion-pumpene er plassert, og endocytic prosesser skje., Mer celleoverflaten mer plass til å romme den molekylære maskiner for exchange. Microvilli er ryggraden som befinner at økt enormt plasma membran overflate og derfor membran protein innhold. Dette er spesielt viktig i de absorptive og secretory celler av epithelia. Microvilli kan øke cellenes overflate opp til 100 ganger sammenlignet med en flat membran. Enterocyte microvilli bære mange enzymer, som ikke er involvert i molekylær exchange, men i fordøyelsen.
Microvilli kan påvirke transduksjon molekylære mekanismer., Deres membraner arbeid som relativt isolert domener med et spesifikt sett av molekyler, som glukose transportører, ion-pumper og-reseptorer. Lengden av microvilli er godt egnet for å utføre lokale transduksjon av molekylære mekanismer uavhengig av cytosol. I tillegg actin filamenter og strukturelle proteiner stillaset danner et tett nettverk av molekyler som liksom filtre eller kontrollerer molekyler kommer eller slutte i det indre av hvert microvellosity. Dette stillaset også kan fungere som en temporær lagring for kalsium.,
høy tetthet av microvilli gjør en fysisk barriere som beskytter mot patogener. Dette er viktig for tarmen epitel. I tillegg er en stor del av membranen som er lagret i microvilli er et reservoar som kan tåle hyperton fornærmelser og hindre brudd på plasma membran.
Stereocilia er spesialisert microvillosities utføre sensoriske funksjoner. Til tross for navnet stereocilium, de er faktisk endret microvillosities forvandlet til sensorisk strukturer, og noen forfattere foreslår navnet estereomicrovellosity i stedet., De kan bli funnet i bitestikkel epitel, og i det indre øret, og arbeidet som mechanoreceptors forstand at de omkringliggende væske bevegelse. Sterocilia av pattedyr indre øret er funnet som en del av Corti ‘ s organ. De er 10 til 50 µm i lengde og inneholder mer enn 3000 actin filament inne. Den lydbølger i luft er forvandlet i væske bølger i øret, som stereocilia fornuft og forvandle seg til elektriske signaler som går via hørselsnerven til hjernen. Stereocilia er som antenne som mottar eksterne signaler, som er transduced i elektriske informasjon.,
Noen microvellosities er spesialisert i å sanse lys. Photorreceptors er lys sensoriske celler som finnes i visual organer som netthinnen. Mobil sensorisk strukturer er faktisk endret cilia eller microvellosities. Fotoreseptorer lager endret microvellosities for light detection kan bli funnet i virvelløse dyr visuelle strukturer. De fotoreseptorer inneholder microvellosities arrangert i den såkalte rhabdomeres. Hver microvellosity inneholder visuelle pigmenter som kan svare for lite lys intensitet og er effektiv ved høye lys intensitet., Microvellosities organisere danner de såkalte rhabdomere, og transduksjon molekylær vei gjøre microvillosity basert lys respons mer fornuftig enn cilium lys sensoriske systemer av vertebrater.
Et stort antall av ekstracellulære blemmer er foreslått å være gitt ut fra microvilli. Tips av enterocyte microvilli er i stand til å frigjøre blemmer til det omkringliggende miljø., Forbindelsen mellom plasma membran og actin-myosin-nettverk kan bidra til å produsere disse blemmer ved å dra membran mot distale del av hver microvellosity, hvor små porsjoner av membranen er forvandlet blemmer. Disse ekstracellulære blemmer inneholder enzymer, og er beriket i alkalisk fosfatase.
Bibliografi
Brown, J W, McKnight C J. 2010. Molekylær modell av microvillar cytoskeleton og organisering av pensel grensen. PLoS One. 5: e940 ![]() .
.
Crawley SW, Mooseker MS, Tyska MJ. 2014. Utformingen av intestinal børste grensen., J Celle Biol. 207: 441-451 ![]() .
.
Fain G L, Hardie R, S Laughlin B. 2010. Phototransduction og utviklingen av fotoreseptorer. Curr Biol. 20: R114-R124 ![]() .
.
Lange K. 2011. Grunnleggende rolle microvilli i de viktigste funksjonene av differensierte celler: Omriss av en universell regulering og signaliserer system på cellen periferien. J Celle Physiol. 226: 896-92
McConnell R E, Higginbotham J N, Shifrin Jr D, Tabb L L, Coffey R J, Tyska M J. 2009. Den enterocyte microvillus er en vesicle-generere organell. J Celle Biol., 185: 1285-129 ![]() .
.
Sauvanet C, Wayt J, Pelaseyed T, Bretscher A. 2015. Struktur, regulering og funksjonelle mangfold av microvilli på apical domenet av epitelceller. Årlig gjennomgang av cellen og utvikling biologi. 31: 593-621.