1. フォーメーション
2. ストラクチャー
3. Function
微絨毛は、通常、いくつかの分化した細胞の自由表面に見られる短い薄い突起であり、上皮細胞でより頻繁に見られる(図1および2)。 それらは、長さ約1-2μm、厚さ約100nmの糸状構造である。 内部では、各微小ビラメントは長軸に平行に配向した数十のアクチンフィラメントを含んでいる。, 微絨毛は通常密で、ブラシの境界線のようにしっかりと詰まっています。 トップビューでは、微じゅう毛は六角形として空間的に組織されています。

図1. 細胞の自由表面に微じゅう毛を示す光学顕微鏡(左の画像)および走査型電子顕微鏡(右の画像)での小腸上皮。
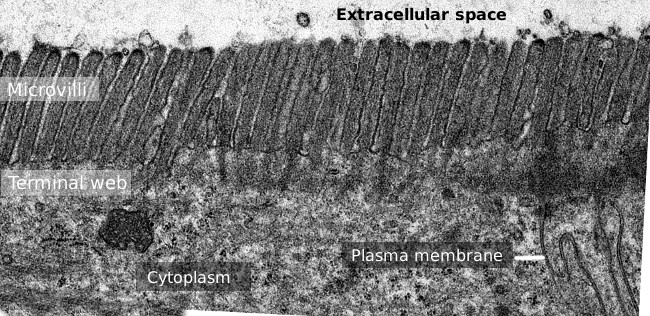
図2. 小腸上皮表面の透過型電子顕微鏡像。, アクチンフィラメントの末端ウェブは、基底から微じゅう毛に観察される暗いバンドである。
微じゅう毛を有する多くの細胞型があるが、腸腸細胞、腎臓の複雑な管上皮および精巣上体のような上皮細胞においてより頻繁に見出される。 微じゅう毛は、嗅覚受容体ニューロン、コルチ受容体細胞の器官、胎盤細胞、および移動している細胞のようないくつかの特殊な感覚細胞にも見られる。, 微じゅう毛は異なる細胞型において同様の外部形態を示すが、それらは明確な分子組成を含み、その内部構造にいくつかの違いを示す。 以下では、小腸上皮の腸細胞の微じゅう毛の特徴を扱っている。
形成
微じゅう毛は、アクチンフィラメントが原形質膜の細胞質ゾル表面に見られる緻密なプレートに関連付けられた後に形成される。 アクチンフィラメントを束に集めて安定化させるためのタンパク質も必要である。 それは順次プロセスのように見えます。, まず、絨毛はアクチンフィラメントの束の形成を促進し、フィラメントの成長を阻害するそれらの遠位端を安定化させるので、絨毛は微じゅう毛の長さを確立するために重要である可能性がある。 エズリンは、あまり豊富ではなく、束のアクチンフィラメントを原形質膜に接続するのに役立つ。 微じゅう毛形成の第二段階は、最終的な長さおよび六角形として細胞表面上の分布に達することを含む。 フィンブリンとスピンはこれらの特徴に寄与するかもしれない。 異なる微じゅう毛のアクチンフィラメント間のプロトカドヘリンによる横方向の接続もこの第二段階の間に起こる。, 微じゅう毛形成中に、大量の膜および膜タンパク質が必要であり、これは激しいエキソサイトーシスによって供給されることに注意することが重要 原形質膜の脱離は,アクチンフィラメント重合の結果である。
構造
微じゅう毛は、アクチン、フィンブリン、ビリン、ミオシン(Myo1A)、カルモジュリン、スペクトリン(非赤血球性)の6つの主要なタンパク質から構成されている(図3)。 マイクロベロシティ骨格は、長軸に平行な30-40アクチンフィラメントの束であり、マイクロベロシティの先端に向かってそれらのマイナス端を有する。, フィンブリンとビリンはアクチンフィラメントを結びつけるが、Myo1Aとカルモジュリンは束と膜の間の接続を作る。 アクチンフィラメントのプラス端は細胞質に延び、そこに異なった微じゅう毛のアクチンフィラメントは互いの間で混合し、六角形パターンの網を形 このウェブはターミナルウェブとして知られており、微絨毛の下の細胞周辺を通って広がっています。 ターミナルウェブには多量の非赤血球性スペクトリンが含まれている。
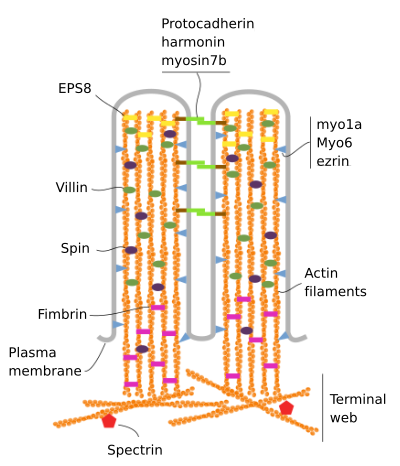
図3., 微じゅう毛の分子組織化(Crowley et al.,2014)
各微小ベロシティは安定であり、移動しないが、そのタンパク質骨格は、微小ベロシティとサイトゾルの間でアクチンおよび他の足場タンパク質を交換することによって永久的に更新されている。 であることを示すバランスと受信および送信をタンパク質 20分ごとにそれぞれのマイクロベロシティの骨格が完成すると推定されている。 微絨毛は、例えば細胞ストレス下で、カルシウム濃度の増加後に完全に解体され、消失することがある。, カルシウムはビリンの挙動を安定化から切断するアクチンフィラメントに変化させる。 微じゅう毛は有糸分裂に入る細胞で消える。 ターミナルウェブもプラスチック構造です。
機能
空洞または外部環境と組織との間の物質の交換は、腸および腎臓上皮などの上皮の主な機能の一つである。 これは、ほとんどの分子トランスポーター、イオンポンプが配置され、エンドサイトーシスプロセスが起こる上皮細胞の頂端(遊離)表面で行われます。, より多くの細胞の表面交換のための分子機械類を収容するより多くのスペース。 微絨毛は、原形質膜表面およびしたがって膜タンパク質content有量を非常に増加させる脊椎様突起である。 これは上皮の吸収性細胞および分泌性細胞のものにおいて特に重要である。 微じゅう毛は平らな膜と比較される100回まで細胞の表面を高めるかもしれません。 腸細胞微絨毛は、分子交換には関与しないが、消化には関与しない多くの酵素を有する。
微じゅう毛は、伝達分子経路に影響を与える可能性があります。, 膜として相対的に孤立したドメイン特異的分子のように、糖トランスポーター、イオンポンプやレーターとして機能している。 微じゅう毛の長さは、サイトゾルとは無関係に分子経路の局所伝達を行うのに適している。 さらに、アクチンフィラメントと構造タンパク質足場は、何らかの形でフィルタリングまたは各微小ベロシティの内部を終了する分子を制御する分子の密なネットワークを形成する。 この足場はまたカルシウムのための一時的な貯蔵として働くかもしれませ,
高密度の微絨毛は、病原体から保護する物理的障壁を作る。 これは腸上皮にとって重要です。 さらに、微じゅう毛に貯蔵された大量の膜は、高張ins辱に耐え、原形質膜の破損を防止することができる貯蔵所である。
立体感症は、感覚機能を実行する特殊な微絨毛である。 Stereociliumという名前にもかかわらず、それらは実際には感覚構造に変換されたマイクロビロシティであり、一部の著者は代わりにestereomicrovellosityという名前を提案している。, それらは、精巣上体上皮および内耳において、周囲の流体の動きを感知する機械受容体として働くことができる。 哺乳類の内耳のステロシリアは、コルティの器官の一部として見出される。 それらは長さが10から50µmで、3000以上のアクチンのフィラメントを中containんでいます。 空気の音波は、耳の流体波に変換され、立体感が感じられ、聴覚神経を介して脳に伝わる電気信号に変換されます。 ステレオシリアは、電気情報に変換される外部信号を受信するアンテナのようなものです。,
いくつかのマイクロベロシティは、光の感知に特化しています。 光受容体は、網膜のような視覚器官に見られる光感覚細胞である。 細胞の感覚構造は、実際には改変された繊毛または微小浮腫である。 光検出のための修正された微小浮腫を有する光受容体は、無脊椎動物の視覚構造に見られる。 これらの光受容体は、いわゆる横紋筋に配置された微小凹凸を含む。 各microvellosityは微光の強度に答えることができ、高い光強度で有効である視覚顔料を含んでいる。, 微小渦はいわゆる横紋を形成する組織し、形質導入分子経路は、脊椎動物の繊毛光感覚系よりも微小渦巻に基づく光応答をより賢明にする。
多数の細胞外小胞が微じゅう毛から放出されることが提案されている。 腸細胞微絨毛の先端は、小胞を周囲の環境に放出することができる。, 原形質膜とアクチン-ミオシンネットワーク間の接続は、膜の小さな部分が小胞内で形質転換される各微小胞の遠位部分に向かって膜をドラッグすることによって、これらの小胞を生成するのに役立つかもしれない。 これらの細胞外小胞は酵素を含み、アルカリホスファターゼに富んでいる。
参考文献
ブラウンJ W、マックナイトC J.2010。 微絨毛細胞骨格の分子モデルおよびブラシ境界の組織。 プロス-ワン 5:e940![]() .
.
クローリーSW、Mooseker MS、ティスカMJ。 2014. 腸のブラシの境界線を整形する。, Jセルバイオール 207:441-451![]() .
.
フェインG L、ハーディR、ラフリンS B.2010。 光伝達と光受容体の進化。 カール-ビオール 20:R114-R124![]() .
.
ランゲK.2011. 分化細胞の主な機能における微じゅう毛の基本的な役割:細胞周辺における普遍的な調節およびシグナル伝達システムの概要。 Jセルフィジオール 226:896-92
McConnell R E,Higginbotham J N,Shifrin Jr D A,Tabb D L,Coffey R J,Tyska M J.2009. エンテロサイトマイクロビルスは、小胞を生成するオルガネラである。 Jセルバイオール, 185:1285-129![]() .
.
Sauvanet C,Wayt J,Pelaseyed T,Bretscher a.2015. 上皮細胞の頂端領域における微じゅう毛の構造、調節、および機能的多様性。 細胞および発生生物学の年次レビュー。 31: 593-621.