1. Muodostelma
2. Rakenne
3. Toiminto
Suolinukan ovat lyhyitä, ohuita ulokkeita yleensä löytyy ilmainen pinnat joitakin erilaisia soluja, useammin epiteelisolujen (Luvut 1 ja 2). Ne ovat noin 1-2 µm pitkiä ja noin 100 nm paksuisia filiformisia rakenteita. Sisällä jokaisessa microvillosityssä on kymmeniä pitkän akselin suuntaisia aktiinifilamentteja., Microvillit ovat yleensä tiheitä ja tiiviisti pakattuja kuin harjaraja. Ylhäältä katsottuna microvillit ovat alueellisesti järjestäytyneitä heksagoneiksi.

Kuva 1. Ohutsuolen epiteelin klo valomikroskoopilla (kuva vasemmalla) ja skannaus elektronimikroskoopilla (kuva oikealla) osoittaa suolinukan vapaa pinta solut.
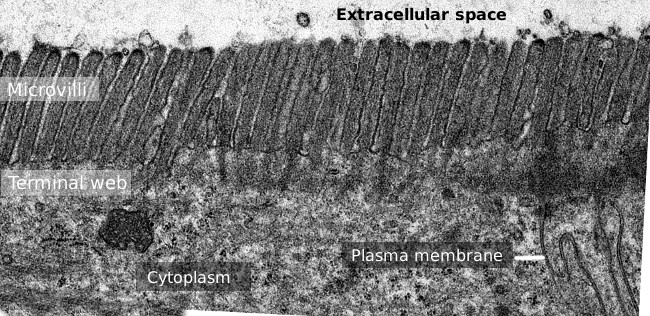
Kuva 2. Siirtoelektronimikroskopia kuva ohutsuolen epiteelin pinta., Terminaali web aktiinifilamenttien on tummempi bändi havaittu basal-insuliinia suolinukan.
On olemassa monia solutyyppejä laakeri suolinukan, mutta ne ovat usein löytyy epiteelisolujen, kuten suolen enterosyyttien, sekava kanava epiteelin munuaisten ja epididymus. Suolinukan löytyy myös joitakin erikoistuneita aisteihin soluja, kuten haju reseptorin neuronien, organ of Corti on reseptorin solut, istukan solut, ja soluja, jotka liikkuvat., Vaikka microvilli osoittaa samanlaista ulkoista morfologiaa eri solutyypeissä, ne sisältävät erillisen molekyylikoostumuksen ja niissä on joitakin eroja niiden sisäisessä rakenteessa. Alla käsitellään ohutsuolen epiteelin enterosyyttien microvilli-ominaisuuksia.
Muodostumisen
Suolinukan on muodostettu sen jälkeen, kun aktiini hehkulankojen saada liittyvät tiheä levy löytyy sytosolisen pinta plasma-kalvo. Tarvitaan myös proteiineja aktiinifilamenttien keräämiseksi nippuihin ja niiden vakauttamiseksi. Se näyttää olevan peräkkäinen prosessi., Ensimmäinen, villin helpottaa muodostumista nippua aktiinifilamenttien ja vakauttaa niiden distaalinen päättyy estämällä hehkulangan kasvaa, niin villin voi olla tärkeää todeta, pituus suolinukan. Vähemmän runsas Ezrin auttaa liittämään nipun aktiinifilamentit plasmakalvostoon. Mikrovillimuodostuksen toiseen vaiheeseen kuuluu lopullisen pituuden saavuttaminen ja jakautuminen solun pinnalla heksagoneina. Fimbrin ja spin voivat edistää näitä ominaisuuksia. Sivusuunnassa yhteydet protocadherins välillä aktiinifilamenttien eri suolinukan myös tässä toisessa vaiheessa., On tärkeää huomata, että aikana suolinukan muodostumista suuri määrä kalvo ja kalvo proteiineja tarvitaan, joka on toimittanut voimakas exocytosis. Plasmakalvon evaginaatiot, yksi jokaista mikrovillositeettia kohti, ovat seurausta aktiinifilamenttipolymeroinnista.
Rakenne
Suolinukan koostuvat 6 tärkeimmät proteiinit: aktiini, fimbrin, villin, myosiini (Myo1A), calmodulin ja spectrin (ei erythrocytic) (Kuva 3). Se microvellosity luuranko on nippu 30-40 aktiinifilamenttien yhdensuuntainen pitkän akselin ja heidän miinus päät kohti kärki microvellosity., Fimbrin ja villin linkki yhteen aktiinifilamenttien, kun taas Myo1A ja calmodulin tehdä yhteyksiä nippu ja kalvo. Plus lopussa aktiinifilamenttien laajentaa solulimaan ja siellä aktiinifilamenttien eri suolinukan sekoittua keskenään ja muodostaa web-kuusikulmainen kuvio. Tämä web tunnetaan terminal web, ja se leviää solun reuna alla microvilli. Terminal web sisältää suuren määrän ei-erytrosyyttistä spektriä.
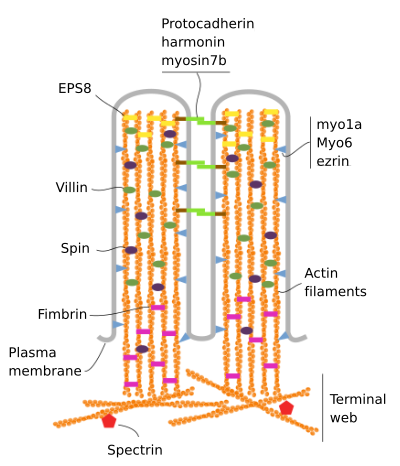
Kuva 3., Microvillin molekyyliorganisaatio (sovitettu Crawley et al., 2014)
Jokainen microvellosity on vakaa ja ei liiku, mutta sen proteiinin luuranko on pysyvä uudistaa vaihtamalla aktiini ja muut telineen proteiinien välillä microvellosity ja sytosolissa. Se tarkoittaa, että saapuvien ja lähtevien proteiinien välillä on tasapaino. On arvioitu, että 20 minuutin välein kunkin microvellosity luuranko on valmis uusitaan. Microvilli voi olla täysin epäjärjestyksessä ja kadota kalsiumin pitoisuuden nousun jälkeen esimerkiksi solustressissä., Kalsium muuttaa villinin käyttäytymistä stabiloitumisesta aktiinifilamenttien katkaisemiseen. Microvilli häviää soluissa, jotka siirtyvät mitoosiin. Terminaaliverkko on myös muovirakenne.
Toiminto
Exchange-aineiden välillä onteloita tai ulkoisen ympäristön ja kudoksissa on yksi päätehtävä epiteelin, kuten suolen ja munuaisten epiteelin. Se on enimmäkseen tehty apikaalisella (ilmainen) pinnan epiteelisolujen, jossa suurin molekyyli transporter, ioni pumput sijaitsevat, ja endocytic prosesseja tapahtua., Mitä enemmän solupintaa, sitä enemmän tilaa vaihtoon mahtuu molekyylikoneistoa. Suolinukan ovat selkärangan-kuten ulokkeita, jotka lisäävät valtavasti solukalvon pinnalla ja siksi kalvo proteiini sisältöä. Tämä on erityisen tärkeää ihosolujen imeytymis-ja erityssoluissa. Microvilli saattaa lisätä solupintaa jopa 100-kertaiseksi verrattuna litteään kalvoon. Enterosyytti microvilli kantaa monia entsyymejä, jotka eivät osallistu molekyylinvaihtoon, vaan ruoansulatukseen.
Microvilli saattaa vaikuttaa transduktion molekyylireitteihin., Kalvot toimivat suhteellisen eristetty verkkotunnuksia eri joukko molekyylejä, kuten glukoosia kuljettajat, ioni -, pumput ja reseptorit. Pituus suolinukan sopii hyvin suorittaa paikallinen transduktio molekyyli polkuja riippumatta sytosolissa. Lisäksi aktiinifilamenttien ja rakenteellisia proteiineja telineen muodostavat tiheän verkoston molekyylejä, jotka jotenkin suodattimia tai ohjaa molekyylien tulossa tai lopettaa sisätilojen kunkin microvellosity. Tämä teline voi myös toimia ajallisena varastona kalsiumille.,
mikrovillien suuri tiheys muodostaa fysikaalisen esteen, joka suojaa taudinaiheuttajilta. Tämä on tärkeää suoliston epiteelille. Lisäksi, suuri määrä kalvo tallennettu suolinukan on säiliö, joka kestää hypertoninen loukkauksia ja estää rikkoutuminen solukalvon.
Stereocilia ovat aistitoimintoja suorittavia erikoistuneita mikrovillositeetteja. Nimestään huolimatta stereocilium, ne ovat itse asiassa muutettu microvillosities muuntaa aistien rakenteet, ja jotkut kirjoittajat ehdottavat, että nimi estereomicrovellosity sijaan., Ne löytyvät lisäkivesten epiteelin, ja sisäkorvan, työskentelee mechanoreceptors mielessä ympäröivän nesteen liikettä. Nisäkkäiden sisäkorvan sterokiliaa tavataan osana Cortin elintä. Niiden pituus on 10-50 µm ja sisällä on yli 3000 aktiinihehkulankaa. Ääniaallot ilmassa muunnetaan nesteen aallot korvaan, joka stereocilia järkeä ja muuntaa sähköiset signaalit, jotka kulkevat kuulohermon kautta aivoihin. Stereocilia on kuin antennit, jotka vastaanottavat ulkoisia signaaleja, jotka transduced sähköinformaatiossa.,
jotkut mikropellot ovat erikoistuneet valon aistimiseen. Fotorekeptorit ovat valoaistisoluja, joita on näköelimissä kuten verkkokalvossa. Soluaistimukset ovat itse asiassa muunneltuja värekarvoja tai mikrovelloja. Photoreceptors laakeri muutettu microvellosities valon havaitseminen löytyy selkärangattomia visuaalisia rakenteita. Nämä fotoreseptorit sisältävät mikrovellositeetteja, jotka on järjestetty niin sanottuihin rabdomereihin. Jokainen mikrovellositeetti sisältää visuaalisia pigmenttejä, jotka voivat vastata alhaisiin valonvoimakkuuksiin ja ovat tehokkaita suurilla valonvoimakkuuksilla., Microvellosities järjestää muodostaen ns rhabdomere, ja transduktio molekyyli-polku saada microvillosity perustuu valon vastaus järkevämpää kuin ripsi valo aistien järjestelmien selkärankaiset.
microvillistä ehdotetaan vapautettavaksi suuri määrä solunulkoisia vesikkeleitä. Enterocyte microvillin kärjet pystyvät vapauttamaan rakkuloita ympäröivään ympäristöön., Yhteys plasman kalvo ja aktiini-myosiini-verkosto voi auttaa tuottamaan nämä rakkulat vetämällä kalvo kohti distaalinen osa jokaisen microvellosity, jossa pieniä osia kalvo on muuttunut rakkulat. Nämä solunulkoiset vesikkelit sisältävät entsyymejä ja rikastuvat emäksisessä fosfataasissa.
bibliografia
Brown J W, McKnight C J. 2010. Molekyylimalli microvillar cytoskeleton ja organisaatio harja rajan. PLoS Yksi. 5: e940 ![]() .
.
Crawley SW, Mooseker MS, Tyska MJ. 2014. Muokkaamalla suoliston harjarajaa., J Cell Biol. 207: 441-451 ![]() .
.
Fain G L, Hardie R, Laughlin S B. 2010. Fototransduktio ja fotoreseptorien kehitys. Curr Biol. 20: R114-R124 ![]() .
.
Lange K. 2011. Perustavanlaatuinen rooli suolinukan tärkein toiminnot eriytetty solut: Outline of an universal säätö-ja merkinantojärjestelmä solun reuna-alueilla. J-Cell Physiol. 226: 896-92
McConnell, R E, Higginbotham J N, Shifrin Jr D, Tabb D L, Coffey R J, Tyska M J. 2009. Enterocyte microvillus on rakkuloita tuottava organelle. J Cell Biol., 185: 1285-129 ![]() .
.
Sauvanet C, Wayt J, Pelasyed T, Bretscher A. 2015. Rakenne, sääntely, ja toiminnallinen monimuotoisuus microvilli on apical domain epiteelisolujen. Solubiologian ja kehitysbiologian vuosikatsaus. 31: 593-621.