1. Formation
2. Struktur
3. Funktion
mikrovilli er korte tynde fremspring, der normalt findes i de frie overflader af nogle differentierede celler, hyppigere i epitelceller (Figur 1 og 2). De er trådformede strukturer på omkring 1 til 2 µm lange og omkring 100 nm tykke. Indvendigt indeholder hver mikrovillositet snesevis af actinfilamenter orienteret parallelt med den lange akse., Microvilli er normalt tæt og tæt pakket som en børstekant. Set ovenfra er mikrovilli rumligt organiseret som sekskanter.

Figur 1. Tyndtarmsepitel ved lysmikroskopi (billede til venstre) og ved scanning af elektronmikroskopi (billede til højre), der viser mikrovilli på cellernes frie overflade.
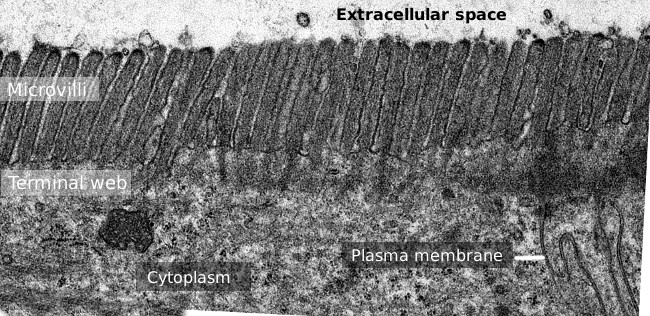
Figur 2. Transmission elektronmikroskopi billede af tyndtarmen epitel overflade., Den terminale bane af actinfilamenter er det mørkere bånd, der observeres basal til mikrovilli.
Der er mange celletyper, der bærer mikrovilli, men de findes hyppigere i epitelceller, som tarmterocytter, indviklet kanalepitel af nyrer og epididymus. Microvilli kan også findes i nogle specialiserede sensoriske celler som olfaktoriske receptor neuroner, organ af Corti receptorceller, placenta celler, og i celler, der bevæger sig., Selvom mikrovilli viser lignende ekstern morfologi i forskellige celletyper, indeholder de særskilt molekylær sammensætning og viser nogle forskelle i deres interne struktur. Nedenfor beskæftiger vi os med funktionerne i mikrovilli af enterocytter i tyndtarmsepitelet.
dannelse
mikrovilli dannes, efter at actinfilamenter er forbundet med en tæt plade, der findes i den cytosoliske overflade af plasmamembranen. Proteiner til indsamling af actinfilamenter i bundter og stabilisering af dem er også nødvendige. Det ser ud til, at det er en sekventiel proces., For det første Letter villin dannelsen af bundter af actinfilamenter og stabiliserer deres distale ender, der hæmmer filamentvæksten, så villin kan være vigtig for at fastslå længden af mikrovilli. E .rin, mindre rigelig, hjælper med at forbinde aktinfilamenterne i bundtet til plasmamembranen. En anden fase af mikrovilli-dannelse inkluderer at nå den endelige længde og fordelingen på celleoverfladen som sekskanter. Fimbrin og spin kan bidrage til disse funktioner. Laterale forbindelser med protocadheriner mellem actinfilamenter af forskellige mikrovilli sker også i denne anden fase., Det er vigtigt at bemærke, at der under mikrovillidannelse er behov for en stor mængde membran-og membranproteiner, som leveres af en intens e .ocytose. Evaginationer af plasmamembran, en pr.
Struktur
Microvilli består af 6 vigtigste proteiner: actin, fimbrin, villin, myosin (Myo1A), calmodulin og spectrin (ikke erythrocytic) (Figur 3). Den microvellosity skeleton er et bundt af 30-40 actin filamenter parallelt med den lange akse og med deres minus ender mod spidsen af microvellosity., Fimbrin og villin forbinder aktinfilamenterne sammen, mens Myo1A og calmodulin skaber forbindelser mellem bundtet og membranen. Den positive ende af actinfilamenter strækker sig ind i cytoplasmaet, og der blandes actinfilamenterne af forskellige mikrovilli mellem hinanden og danner en bane med et sekskantet mønster. Denne webeb er kendt som terminal webeb, og det spredes gennem cellens periferi under mikrovilli. Terminal webeb indeholder store mængder af ikke-erythrocytisk spectrin.
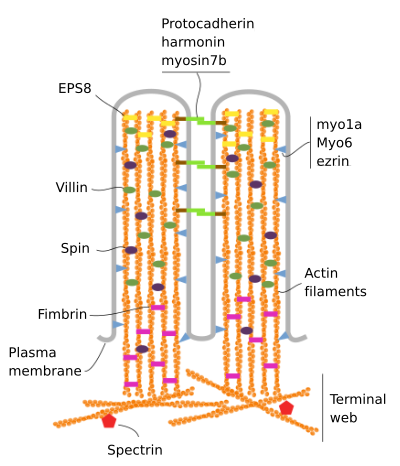
Figur 3., Molekylær organisation af microvilli (tilpasset fra CRA .ley et al., 2014)
hver mikrovellositet er stabil og bevæger sig ikke, men dets proteinskelet er i permanent fornyelse ved at udveksle actin og andre stilladsproteiner mellem mikrovellositeten og cytosolen. Det betyder, at der er en balance mellem indgående og udgående proteiner. Det anslås, at hvert 20. minut er skeletet af hver mikrovellositet afsluttet fornyet. Mikrovilli kan være fuldstændig uorganiseret og forsvinde efter en stigning i calciumkoncentration, for eksempel under cellespænding., Calcium ændrer Villins opførsel, fra stabilisering til adskillelse af actinfilamenter. Mikrovilli forsvinder i celler, der kommer ind i mitose. Terminalbanen er også en plastikstruktur.
funktion
udveksling af stoffer mellem hulrum eller ydre miljø og væv er en hovedfunktion af epitel, såsom tarm og nyreepitel. Det gøres for det meste ved den apikale (frie) overflade af epithelcellerne, hvor de fleste molekylære transporter, ionpumper er placeret, og endocytiske processer sker., Jo mere celleoverflade jo mere plads til at rumme det molekylære maskiner til udvekslingen. Mikrovilli er rygsøjlelignende fremspring, der øger plasmamembranoverfladen enormt og derfor membranproteinindholdet. Dette er særligt vigtigt i de absorberende og sekretoriske celler af epithelia. Mikrovilli kan øge celleoverfladen op til 100 gange sammenlignet med en flad membran. Enterocytmikrovilli bærer mange en .ymer, som ikke er involveret i Molekylær udveksling, men i fordøjelsen.
mikrovilli kan påvirke transduktionsmolekylære veje., Deres membraner fungerer som relativt isolerede domæner med et særskilt sæt molekyler, som glukosetransportører, ionpumper og receptorer. Længden af mikrovilli er velegnet til at udføre lokal transduktion af de molekylære veje uafhængigt af cytosolen. Derudover danner actinfilamenterne og strukturelle proteiner stillads et tæt netværk af molekyler, der på en eller anden måde filtrerer eller styrer molekylerne, der kommer eller afslutter det indre af hver mikrovellositet. Dette stillads kan også fungere som en tidsmæssig opbevaring af calcium.,
den høje densitet af mikrovilli gør en fysisk barriere, der beskytter mod patogener. Dette er vigtigt for tarmepitel. Derudover er den store mængde membran, der opbevares i mikrovilli, et reservoir, der kan modstå hypertoniske fornærmelser og forhindre brud på plasmamembranen.
Stereocilia er specialiserede mikrovillositeter, der udfører sensoriske funktioner. På trods af navnet stereocilium er de faktisk modificerede mikrovillositeter omdannet til sensoriske strukturer, og nogle forfattere foreslår i stedet navnet estereomicrovellosity., De kan findes i epididymisepitelet og i det indre øre, der fungerer som mekanoreceptorer, der fornemmer den omgivende væskebevægelse. Sterocilia af pattedyrets indre øre findes som en del af Cortis organ. De er 10 til 50 µm i længden og indeholder mere end 3000 actin filament indeni. Lydbølgerne i luften omdannes i væskebølger i øret, som stereocilia fornemmer og omdanner til elektriske signaler, der bevæger sig via hørenerven til hjernen. Stereocilia er som antenner, der modtager eksterne signaler, som transduceres i elektrisk information.,
nogle mikrovellositeter er specialiserede i at registrere lys. Fotorreceptorer er lette sensoriske celler, der findes i visuelle organer som nethinden. De cellulære sensoriske strukturer er faktisk modificerede cilia eller mikrovellositeter. Fotoreceptorer, der bærer modificerede mikrovellositeter til lysdetektion, findes i hvirvelløse visuelle strukturer. Disse fotoreceptorer indeholder mikrovellositeter arrangeret i de såkaldte rhabdomerer. Hver mikrovellositet indeholder visuelle pigmenter, der kan reagere på lav lysintensiteter og er effektive ved høje lysintensiteter., Microvellosities organisere danner den såkaldte rhabdomere, og transduktion molekylær vej gøre microvillosity baseret lette svar er mere fornuftig end cilium lys sensoriske systemer af hvirveldyr.
et stort antal ekstracellulære vesikler foreslås frigivet fra mikrovilli. Spidserne af enterocytmikrovilli er i stand til at frigive vesikler til det omgivende miljø., Forbindelsen mellem plasmamembran og actin-myosin netværk kan bidrage til at producere disse vesikler ved at trække membranen mod den distale del af hver mikrovellositet, hvor små dele af membranen transformeres i vesikler. Disse ekstracellulære vesikler indeholder en .ymer og er beriget med alkalisk fosfatase.
bibliografi
bro .n J,, McKnight C J. 2010. Molekylær model af mikrovillar cytoskelettet og organisering af børsten grænsen. PLoS One. 5: e940 ![]() .
.
Crawley SW, Mooseker MS, Tyska MJ. 2014. Udformning af tarmbørste grænsen., J Cell Biol. 207: 441-451 ![]() . Fain G L, Hardie R, Laughlin s B. 2010. Fototransduktion og udviklingen af fotoreceptorer. Curr Biol. 20: R114-R124
. Fain G L, Hardie R, Laughlin s B. 2010. Fototransduktion og udviklingen af fotoreceptorer. Curr Biol. 20: R114-R124 ![]() .
.
Lange K. 2011. Grundlæggende rolle microvilli i de vigtigste funktioner i differentierede celler: omrids af et universelt regulerings-og signalsystem ved cellens periferi. J Cellefysiol. 226: 896-92
McConnell R E, Higginbotham J N, Shifrin Jr D A, Tabb D L, Coffey R J, Tyska M J. 2009. Enterocyt microvillus er en vesikel-genererende organelle. J Cell Biol., 185: 1285-129 ![]() .
.
Sauvanet C, Wayt J, Pelaseyed T, Bretscher A. 2015. Struktur, regulering og funktionel mangfoldighed af mikrovilli på det apikale domæne af epitelceller. Årlig gennemgang af celle-og udviklingsbiologi. 31: 593-621.