1. Formace
2. Struktura
3. Funkce
Mikroklků jsou krátké, tenké výčnělky obvykle nalézt ve volné povrchy některých diferencovaných buněk, častější v epitelových buněk (obrázky 1 a 2). Jsou to filiformní struktury o délce asi 1 až 2 µm a tloušťce kolem 100 nm. Uvnitř každá mikrovilozita obsahuje desítky aktinových vláken orientovaných rovnoběžně s dlouhou osou., Mikrovily jsou obvykle husté a pevně zabalené jako okraj štětce. V horním pohledu jsou mikrovily prostorově organizovány jako šestiúhelníky.

Obrázek 1. Malé střevní epitel na světelné mikroskopie (obrázek vlevo) a skenovací elektronové mikroskopie (obrázek vpravo) ukazuje, microvilli na volném povrchu buněk.
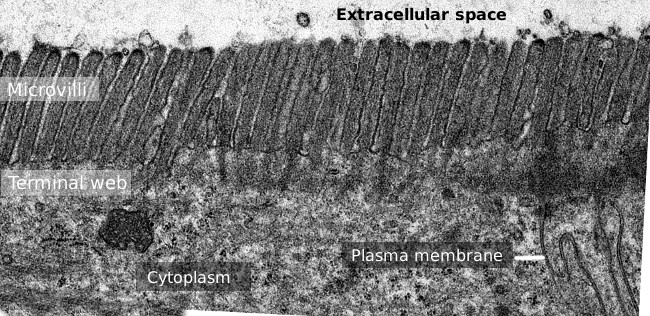
Obrázek 2. Transmisní elektronová mikroskopie obraz povrchu epitelu tenkého střeva., Koncová síť aktinových vláken je tmavší pás pozorovaný bazální až mikrovilli.
Existuje mnoho typů buněk, opatřený mikroklky, ale oni jsou více často nalezený v epitelových buněk, jako střevní enterocyty, spletitý potrubí epitel ledvin a epididymus. Mikrovily lze také nalézt v některých specializovaných senzorických buňkách, jako jsou neurony čichového receptoru, orgán buněk receptoru Corti, placentární buňky a buňky, které se pohybují., Ačkoli mikrovily vykazují podobnou vnější morfologii v různých typech buněk, obsahují odlišné molekulární složení a vykazují určité rozdíly ve své vnitřní struktuře. Níže se zabýváme vlastnostmi mikrovilů enterocytů epitelu tenkého střeva.
Tvorbu
Mikroklků jsou tvořeny po aktinová filamenta dostat spojena s hustou deska nalezena v cytosolu povrchu plazmatické membrány. Proteiny pro shromažďování aktinových vláken ve svazcích a jejich stabilizaci jsou také potřebné. Vypadá to, že se jedná o postupný proces., První, villin usnadňuje tvorbu svazky aktinových vláken a stabilizuje jejich distální konce inhibiční vlákna rostou, takže villin může být důležité stanovit délku mikroklků. Ezrin, méně hojný, pomáhá spojit aktinové vlákna svazku s plazmatickou membránou. Druhá fáze tvorby mikrovilů zahrnuje dosažení konečné délky a rozdělení na buněčný povrch jako šestiúhelníky. Fimbrin a spin mohou přispět k těmto funkcím. Boční připojení protocadherins mezi aktinová filamenta různých mikroklků se také stát, během této druhé fáze., Je důležité si uvědomit, že během tvorby mikrovilů je zapotřebí velké množství membránových a membránových proteinů, které jsou dodávány intenzivní exocytózou. Evaginace plazmatické membrány, jedna na každou mikrovilozitu, je důsledkem polymerace aktinových vláken.
Struktura
Mikroklků jsou vyrobeny z 6 hlavní bílkoviny: aktin, fimbrin, villin, myosin (Myo1A), calmodulin a spectrin (non erytrocytární) (Obrázek 3). Kostra mikrovelozity je svazek 30-40 aktinových vláken rovnoběžných s dlouhou osou a s jejich mínusovými konci směrem ke špičce mikrovelozity., Fimbrin a villin spojují aktinová vlákna, zatímco Myo1A a calmodulin vytvářejí spojení mezi svazkem a membránou. Na plus konci aktinová filamenta rozšířit do cytoplazmy a tam aktinová filamenta různých mikroklků prolínají mezi sebou a tvořit web s šestihranných vzor. Tento web je známý jako terminální web a šíří se přes buněčný obvod pod mikrovilli. Terminální síť obsahuje velké množství non-erytrocytického spektrinu.
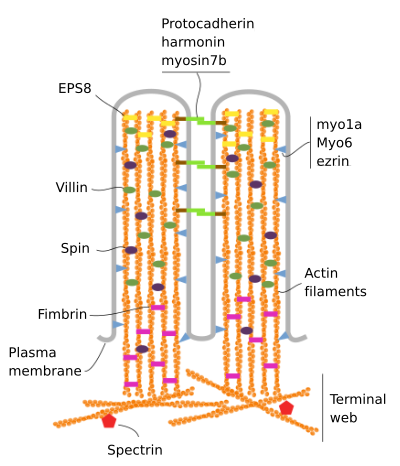
obrázek 3., Molekulární organizace mikrovilli (převzato z Crawley et al., 2014)
Každý microvellosity je stabilní a nepohybuje, ale jeho protein kostra je v trvalé obnovení výměnou aktin a jiné lešení proteinů mezi microvellosity a cytosolu. To znamená, že existuje rovnováha mezi příchozími a odchozími bílkovinami. Odhaduje se, že každých 20 minut je kostra každé mikrovelozity dokončena. Mikrovily mohou být zcela dezorganizované a zmizí po zvýšení koncentrace vápníku, například při buněčném stresu., Vápník mění chování villinů, od stabilizace po oddělení aktinových vláken. Mikrovily zmizí v buňkách vstupujících do mitózy. Svorkovnice je také plastová konstrukce.
Funkce
Výměna látek mezi dutiny nebo na vnější prostředí a tkání, je jedním z hlavních funkce epitelu, jako jsou střeva a ledvin epitelu. Většinou se provádí na apikálním (volném) povrchu epiteliálních buněk, kde se nachází většina molekulárního transportéru, iontových čerpadel a dochází k endocytickým procesům., Čím více buněčného povrchu, tím více prostoru pro umístění molekulárních strojů pro výměnu. Mikrovily jsou páteřní výčnělky, které enormně zvyšují povrch plazmatické membrány, a tedy obsah membránových bílkovin. To je zvláště důležité v těch absorpčních a sekrečních buňkách epitelu. Mikrovily mohou zvýšit buněčný povrch až 100krát ve srovnání s plochou membránou. Enterocytové mikrovily nesou mnoho enzymů, které se nepodílejí na molekulární výměně, ale na trávení.
Mikrovily mohou ovlivňovat transdukční molekulární dráhy., Jejich membrány fungují jako relativně izolované domény s odlišnou sadou molekul, jako jsou transportéry glukózy, iontové pumpy a receptory. Délka mikrovilů je vhodná pro provádění lokální transdukce molekulárních cest nezávisle na cytosolu. Kromě toho, aktinová filamenta a strukturální proteiny lešení tvoří hustou síť molekul, které nějak filtry nebo kontroly molekuly pocházející nebo ukončení každého interiéru microvellosity. Toto lešení může také fungovat jako časové úložiště vápníku.,
vysoká hustota mikrovilů vytváří fyzickou bariéru, která chrání před patogeny. To je důležité pro střevní epitel. Kromě toho, velké množství membrány uložené v mikroklků je nádrž, která vydrží hypertonický urážky a zabránit poškození plazmatické membrány.
Stereocilia jsou specializované mikrovillosity provádějící senzorické funkce. Přes název stereocilium jsou ve skutečnosti modifikované mikrovilozity transformované do senzorických struktur a někteří autoři místo toho navrhují název estereomicrovellosity., Mohou se nalézt v epitelu epididymis a ve vnitřním uchu, které pracují jako mechanoreceptory, které snímají pohyb okolní tekutiny. Sterocilia vnitřního ucha savců se nachází jako součást Cortiho orgánu. Mají délku 10 až 50 µm a uvnitř obsahují více než 3000 aktinových vláken. Zvukové vlny ze vzduchu se mění v kapalinu vlny v uchu, které stereocilia smysl a přeměnit na elektrické signály, které se šíří prostřednictvím sluchového nervu do mozku. Stereocilia jsou jako antény, které přijímají externí signály, které jsou přenášeny v elektrických informacích.,
některé mikrovellosity se specializují na snímání světla. Fotoreceptory jsou světelné senzorické buňky nalezené ve zrakových orgánech, jako je sítnice. Buněčné senzorické struktury jsou ve skutečnosti modifikované řasy nebo mikrovellosity. Fotoreceptory nesoucí modifikované mikrovellosity pro detekci světla lze nalézt ve vizuálních strukturách Bezobratlých. Tyto fotoreceptory obsahují mikrovellosity uspořádané v tzv. Každá mikrovelozita obsahuje vizuální pigmenty, které mohou reagovat na nízké intenzity světla a jsou účinné při vysokých intenzitách světla., Microvellosities organizovat tvoří tzv. rhabdomere, a transdukce molekulární dráhy, aby microvillosity na bázi světelné odezvy rozumnější než cilium světlo smyslových systémů obratlovců.
z mikrovilů se navrhuje uvolnit velké množství extracelulárních váčků. Špičky enterocytových mikrovilů jsou schopny uvolňovat vezikuly do okolního prostředí., Spojení mezi plazmatickou membránou a actin-myosinovou sítí může pomoci vytvořit tyto vezikuly přetažením membrány směrem k distální části každé mikrovelozity, kde se malé části membrány transformují ve vezikulech. Tyto extracelulární váčky obsahují enzymy a jsou obohaceny alkalickou fosfatázou.
bibliografie
Brown J W, McKnight C J. 2010. Molekulární model mikrovilárního cytoskeletu a organizace okraje štětce. PLoS Jedna. 5: e940 ![]() .
.
Crawley SW, Mooseker MS, Tyska MJ. 2014. Tvarování okraje střevního kartáče., J Cell Biol. 207: 441-451 ![]() .
.
Fain g L, Hardie R, Laughlin s B. 2010. Fototransdukce a vývoj fotoreceptorů. Curr Biol. 20: R114-R124 ![]() .
.
Lange k. 2011. Základní úloha mikrovilli v hlavních funkcích diferencovaných buněk: obrys univerzálního regulačního a signalizačního systému na obvodu buněk. J Cell Physiol. 226: 896-92
McConnell R E, Higginbotham J N, Shifrin Jr D A, Tabb D L, Coffey R J, Tyska M J.2009. Enterocyt microvillus je organela vytvářející vezikuly. J Cell Biol., 185: 1285-129 ![]() .
.
Sauvanet C, Wayt J, Pelaseyed T, Bretscher a. 2015. Struktura, regulace a funkční rozmanitost mikrovilli na apikální doméně epiteliálních buněk. Roční přehled buněčné a vývojové biologie. 31: 593-621.